1.Introduction.
Photosynthesis in all algae and higher plants takes place in the presence of chlorophyll a; the exception are photosynthetic bacteria, which contain pigments with a slightly different structure than chlorophyll a. Other varieties of chlorophyll also occur in plants.
Higher plants and green algae contain chlorophyll a and b, while cyanobacteria and some red algae contain only chlorophyll a. Other red algae species contain chlorophyll a and a small amount of chlorophyll d, and brown algae and diatoms have both chlorophyll aand chlorophyll c.
In addition, other pigments participate in photosynthesis, such as carotenoids, and in red algae and cyanobacteria, phycobilin pigments (phycoerythrin and phycocyanin). Table 1 lists pigments found in various photosynthetic organisms, including their color and primary absorption maximum. Figure 1 shows the basic chemical structures of some photosynthetic pigments.
Fig. 1. Structural formulas of chlorophylls.
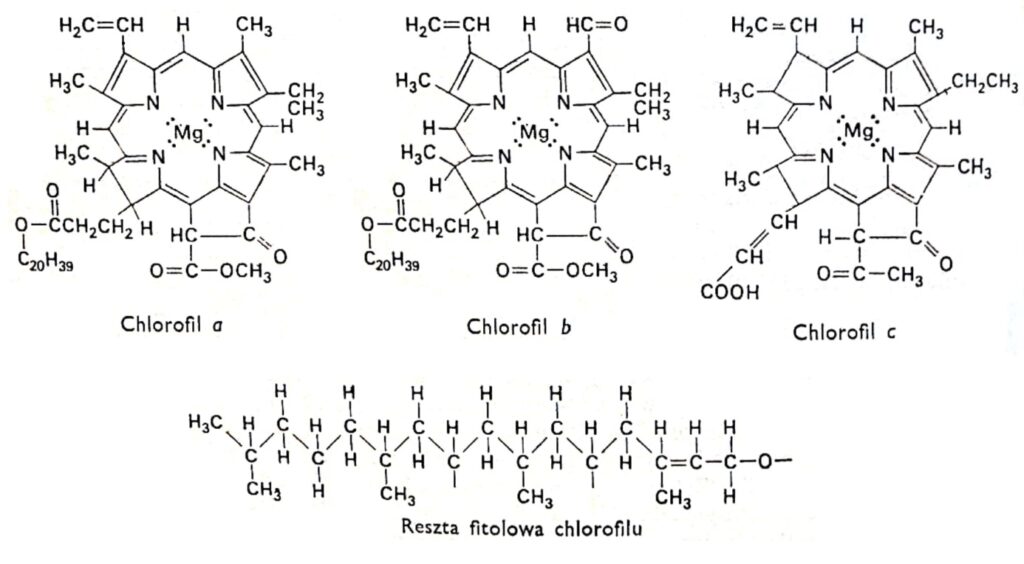
Table 1. Properties of photosynthetic pigments.
Dye | Color | Main absorption peaks in the visible range (nm) |
| Chlorophyll a | blue-green | 430 (blue-violet) 660 (red) |
| Chlorophyll b | green | 455 (blue) 640 (orange-red) |
| Phycocyanins | blue | 560-660 (yellow to orange-red) |
| Phycoerythrins | red | 550-570 (green to yellow) |
| Carotenoids | Yellow, orange-red | 430-490 (purple to blue-green) |
2. The structure of chlorophyll.
All varieties of chlorophyll have the same porphyrin structure, composed of four pyrrole rings linked to magnesium via nitrogen atoms. Furthermore, the chlorophyll molecule contains a fifth ring composed entirely of carbon atoms and linked to the long-chain alcohol phytol, typical of most chlorophylls, with the exception of chlorophyll c. Chlorophylls differ in the structure of the side chains attached to the pyrrole rings. The difference between chlorophyll a and b is that the third ring of chlorophyll a contains a methyl group, while in chlorophyll b it contains an aldehyde group. The aldehyde group makes chlorophyll b more hydrophilic (soluble in 92% methyl alcohol) than chlorophyll a (more soluble in ether). The vast majority of plants contain two or three times more chlorophyll a than chlorophyll b. The structure of the chlorophyll molecule was determined at the beginning of this century by the German researchers R. Wilstatter and A. Stoll and the famous organic chemist Hans Fischer.
Chlorophylls a and b absorb primarily blue-violet and red light, and
only slightly green light. For chlorophyll a, the maximum absorption occurs in the violet range, i.e., at shorter wavelengths than for chlorophyll b; whereas in the red range, the maximum is shifted toward longer wavelengths. "The position of the absorption maximum depends on the solvent in which the dyes are dissolved, i.e., on the type of solvent molecules with which the dye molecules are associated. For example, in a living plant, the main absorption peak of chlorophyll a in the red range is at 683 nm, which is shifted in relation to the peak at 660 nm for this chlorophyll dissolved in ethyl ether. For chlorophyll a, an additional absorption peak at 672 nm was also found. It is therefore assumed that this is the result of slightly changed environmental conditions. Graphs showing the dependence of light absorption on different wavelengths are called absorption spectra. The absorption spectrum is determined using a spectrophotometer, i.e., an instrument equipped with a light source that emits waves of the desired wavelengths; a prism or a diffraction grating that serves to disperse the light; mirrors that reflect selected wavelengths and direct them onto the sample being tested, and A detector used to detect the amount of light absorbed by the sample being tested. The irradiated chlorophyll solution fluoresces strongly, emitting intense red light. Because most of the energy that causes fluorescence in living plants is used in photosynthesis, they do not fluoresce visibly.
3. Chlorophyll synthesis.
Chlorophyll synthesis is dependent on a number of factors, including genetic characteristics, light, and the availability of certain mineral elements. In certain varieties of corn and sunflowers, some germinating seeds produce seedlings lacking chlorophyll. This is an innate recessive trait. These shoots, called albinos, die when the seed's nutritional reserves are exhausted. However, if these plants are supplied with sugars as an energy source, they can continue to grow until almost maturity.
Light is essential for chlorophyll synthesis in almost all angiosperms, although conifers, mosses, some ferns, and many algae can also produce this pigment in the dark. Angiosperms growing in the dark synthesize small amounts of a green pigment similar to chlorophyll, but its content is too low to impart a green color to these plants. Upon exposure to light, this pigment disappears, replaced by chlorophyll a, the amount of which increases with time. This disappearing pigment is called protochlorophyll. Chemically, it differs from chlorophyll a only by the absence of two hydrogen atoms in the molecule. Furthermore, it absorbs blue and red light similarly, exhibiting absorption maxima at wavelengths of 450 nm and 650 nm. These wavelengths of light are most effective in the conversion of protochlorophyll to chlorophyll a. However, the origin of chlorophyll b remains unexplained.
It appears that a number of mineral elements directly participate in chlorophyll synthesis, as their deficiency causes discoloration, or chlorosis, in plants. Chlorophyll pyrrole rings are formed by the prior combination of glycine with succinic acid. Between the reaction of these two compounds and the final formation of the complex compound chlorophyll, a series of intermediate reactions catalyzed by specific enzymes occur. The reaction between glycine and succinyl-CoA is dependent on the presence of iron as a coenzyme, and a deficiency of this element causes rapid chlorosis in young, developing leaves. Deficiencies of manganese, potassium, zinc, and copper also cause chlorosis, but the role of these elements is not yet understood. They presumably activate certain enzymes involved in chlorophyll synthesis. Manganese deficiency leads to the destruction of the internal lamellar structure of chloroplasts and may be the reason for the inhibition of the synthesis of chlorophyll stabilizing the lamellae.
However, a lack of nitrogen and magnesium causes yellowing of plants. The role of these elements is understandable, as they are the basic structural components of chlorophyll. This does not rule out their more indirect functions, for example, magnesium likely acts as a coenzyme in one or more steps of chlorophyll synthesis.
4. Structure and Synthesis of Carotenoid Pigments.
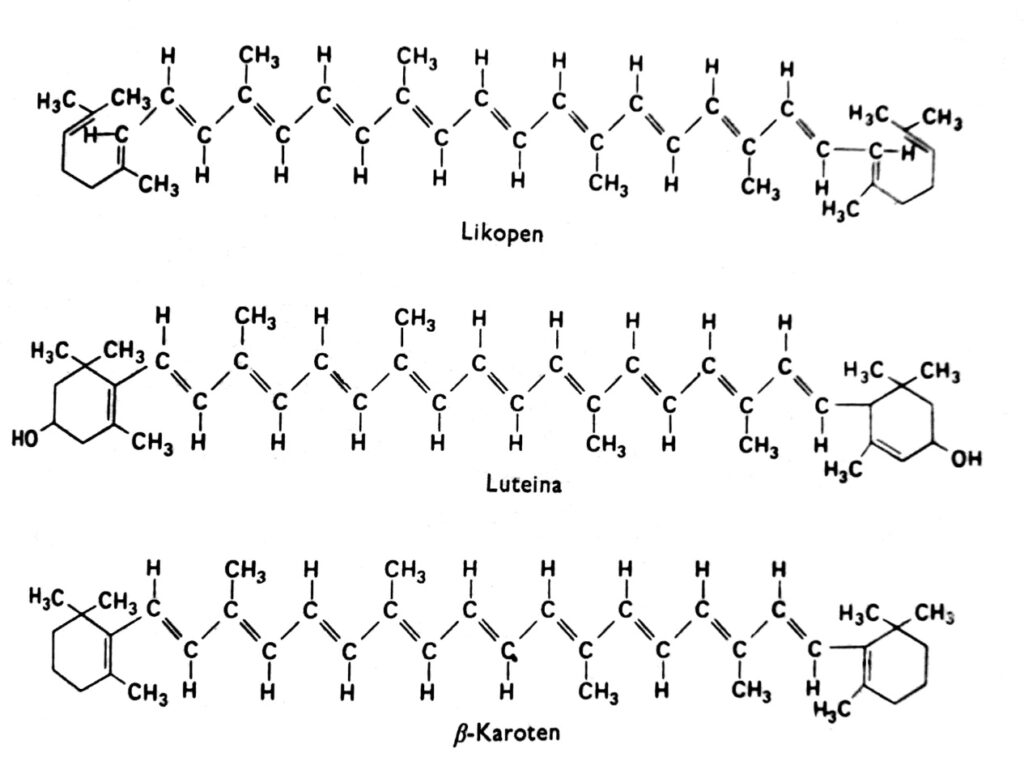
Carotenoids are a group of pigments that, in higher plants, are generally red, orange, yellow, or brown and are found in chloroplasts in combination with chlorophyll. They are also sometimes found in other plastids, such as the chromoplasts of carrot and tomato roots, and in the chromoplasts of numerous yellow flowers. The yellow color of carotenoids is visible in angiosperms growing in the dark and lacking chlorophyll (indicating that light is not necessary for their accumulation) and in autumn in the leaves of many species, where chlorophyll disappears. Indeed, carotenoids can also exhibit other colors, such as green in avocados, pink in shrimp, or almost ink-black in cephalopods. These pigments occur in all major groups of plants and animals, but our discussion will focus primarily on those found in chloroplasts in combination with chlorophyll. Carotenoids can be divided into two chemical groups: carotenes and xanthophylls. Within each group, there are several distinct types of these compounds, whose molecules are composed of 40 carbon atoms. Carotenoids, the most important representative of which is β-carotene, are hydrocarbons (Fig. 2). Xanthophylls, such as lutein and zeaxanthin, have oxygen atoms attached to their terminal rings. Both xanthophylls and carotenes are synthesized in plants from active acetate, i.e., acetyl-CoA, via mevalonic acid.
Fig. 2. Structural formulas of carotenoids.
Almost none of the carotenoids dissolve in water, but they do dissolve in many organic solvents, such as acetone and ether. Xanthophylls, having additional oxygen atoms, are more hydrophilic than carotenes. In solution, most carotenoids absorb primarily violet and blue light in the 400-500 nm range. Their absorption spectra depend on the degree of double bond saturation, and therefore may be slightly different in intact chloroplasts. There is evidence that in intact cells, the absorption maxima of some carotenoids at 500 nm are shifted to 450 nm after extraction of these pigments with an organic solvent.
5. Functions of carotenoids.
Carotenoids play an important role in photosynthesis, probably for two reasons:
1. They perform a protective function, protecting chlorophyll from the damaging effects of light
and oxygen. This destructive phenomenon is called photooxidation. It is often observed that an alcoholic solution of chlorophyll discolors under the influence of light, which is accompanied by oxygen consumption. Presumably, a similar reaction would normally occur in plants if carotenoids did not fulfill their protective function. The most convincing evidence comes from observations of certain bacterial mutants. Because they do not contain carotenoid pigments, photooxidation occurs very readily in them. Furthermore, albino sunflower mutants are known that have lost the ability to synthesize carotenoids but still produce trace amounts of chlorophyll. In this case, increasing light intensity triggers additional photooxidation of chlorophyll, and the rate of this process is equal to its synthesis. The mechanism by which carotenoids protect chlorophyll from oxidation remains unclear. However, it is believed that carotenoids are photooxidized by light, which could decompose chlorophyll.
2. Another likely function of carotenoids, and especially xanthophylls, is their involvement in the absorption of light at wavelengths active in photosynthesis. There is indisputable evidence that some carotenoids, such as fucoxanthin (also called fucoxanthol), found in brown algae (e.g., fucus), actively participate in photosynthesis. This evidence was obtained by comparing the absorption spectrum of fucoxanthin with the so-called functional spectrum, or the spectrum of photosynthetic activity. The photosynthetic functional spectrum is a graph illustrating the efficiency of photosynthesis as a function of the wavelengths of light that trigger this process. It has been found that in green plants and higher plants, light absorbed by some carotenoids is also used in photosynthesis. In such cases, the maxima of the functional spectrum fall in the red and blue light ranges, where chlorophyll absorbs most strongly. However, photosynthesis still occurs using blue-green light, presumably because some carotenoids are active in this range. The most compelling evidence that carotenoids participate in photosynthesis is their ability to transfer excited energy directly to chlorophyll a. When a plant is irradiated with light at wavelengths absorbed almost exclusively by carotenoids, red fluorescence from chlorophyll a is observed. It is hypothesized that the excitation energy, sometimes released as fluorescence, is typically used as the driving force for photosynthesis.
6. Phycobilins.
The final group of photosynthetic pigments are phycobilins, found in cyanobacteria
and red algae. These pigments are classified into two groups: red (phycoerythrins) and blue (phycocyanins). Like chlorophyll, they are composed of four pyrrole rings. However, phycobilins contain an open pyrrole ring system and neither magnesium nor a phytol moiety. In living cells, they occur in association with proteins and are water-soluble. Both types of pigments absorb light, using it for photosynthesis. In red algae, the highest photosynthetic efficiency occurs in the green light range, because in this range, phycoerythrin, unlike chlorophyll, absorbs light most intensely. Like carotenoids, phycobilins transfer excited energy to chlorophyll a and can cause it to fluoresce. This mode of energy transfer indicates that phycobilins actively participate in photosynthesis by capturing photons, the energy of which is typically used by chlorophyll a as the driving force for photosynthesis. Pigments such as carotenoids and phycobilins, which transfer energy to a suitable acceptor, e.g., chlorophyll a, are called accessory pigments. The group of accessory pigments also includes chlorophylls b, c , and d, because they absorb light energy, which in turn can be transferred to chlorophyll a.
Literature:
-Franck B. Salibury, Cleon Ross "Plant Physiology" PWRiL 1975.
-Otis F. Daniel G. Curtis Clark "Introduction to Plant Physiology" PWRiL 1958.
-Zurzycki J. Michniewicz M. "Plant Physiology" PWRiL 1979.
-Richter G. "Metabolic processes in plants" PWN 1975.