1. Introdução.
A fotossíntese em todas as algas e plantas superiores ocorre na presença de clorofila a; a exceção são as bactérias fotossintéticas, que contêm pigmentos com uma estrutura ligeiramente diferente da clorofila a. Outras variedades de clorofila também ocorrem em plantas.
As plantas superiores e as algas verdes contêm clorofila a e b, enquanto as cianobactérias e algumas algas vermelhas contêm apenas clorofila a. Outras espécies de algas vermelhas contêm clorofila a e uma pequena quantidade de clorofila d, e as algas pardas e as diatomáceas possuem clorofila ae clorofila c.
Além disso, outros pigmentos participam da fotossíntese, como os carotenoides e, em algas vermelhas e cianobactérias, os pigmentos ficobilínicos (ficoeritrina e ficocianina). A Tabela 1 lista os pigmentos encontrados em diversos organismos fotossintéticos, incluindo sua cor e máximo de absorção primário. A Figura 1 mostra as estruturas químicas básicas de alguns pigmentos fotossintéticos.
Figura 1. Fórmulas estruturais das clorofilas.
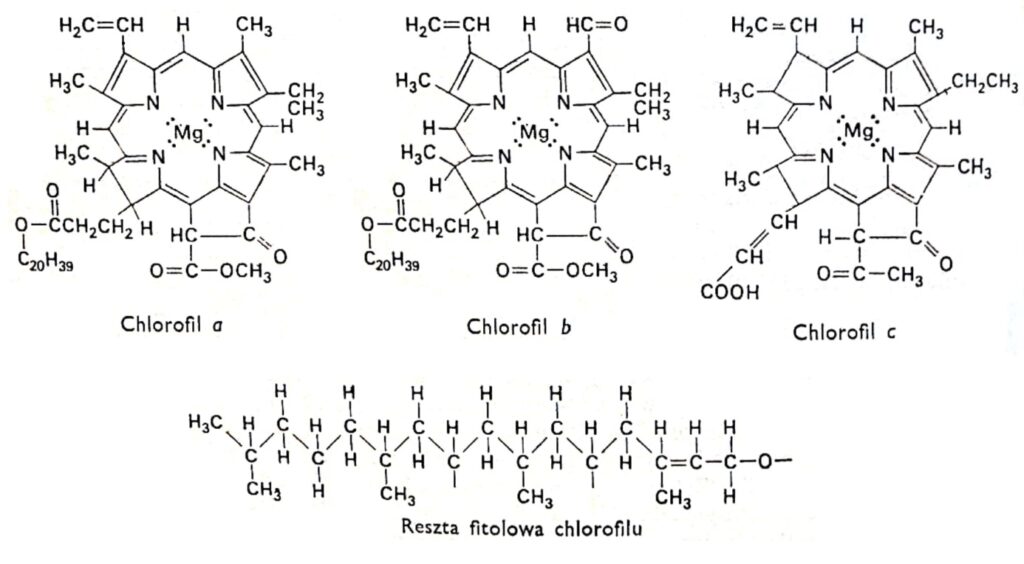
Tabela 1. Propriedades dos pigmentos fotossintéticos.
Tingir | Cor | Principais picos de absorção na faixa visível (nm) |
| Clorofila a | azul-esverdeado | 430 (azul-violeta) 660 (vermelho) |
| Clorofila b | verde | 455 (azul) 640 (vermelho-alaranjado) |
| Ficocianinas | azul | 560-660 (amarelo a vermelho-alaranjado) |
| Ficoeritrinas | vermelho | 550-570 (verde a amarelo) |
| Carotenoides | Amarelo, vermelho-alaranjado | 430-490 (roxo a verde-azulado) |
2. A estrutura da clorofila.
Todas as variedades de clorofila possuem a mesma estrutura porfirínica, composta por quatro anéis pirrólicos ligados ao magnésio por meio de átomos de nitrogênio. Além disso, a molécula de clorofila contém um quinto anel composto inteiramente de átomos de carbono e ligado ao álcool de cadeia longa fitol, típico da maioria das clorofilas, com exceção da clorofila c. As clorofilas diferem na estrutura das cadeias laterais ligadas aos anéis pirrólicos. A diferença entre a clorofila a e a b reside no fato de que o terceiro anel da clorofila a contém um grupo metil, enquanto na clorofila b contém um grupo aldeído. O grupo aldeído torna a clorofila b mais hidrofílica (solúvel em metanol a 92%) do que a clorofila a (mais solúvel em éter). A grande maioria das plantas contém duas ou três vezes mais clorofila a do que clorofila b. A estrutura da molécula de clorofila foi determinada no início deste século pelos pesquisadores alemães R. Wilstatter e A. Stoll e pelo famoso químico orgânico Hans Fischer.
As clorofilas a e b absorvem principalmente luz azul-violeta e vermelha, e
apenas ligeiramente luz verde. Para a clorofila a, a absorção máxima ocorre na faixa do violeta, ou seja, em comprimentos de onda mais curtos do que para a clorofila b; enquanto que na faixa do vermelho, o máximo é deslocado para comprimentos de onda mais longos. A posição do máximo de absorção depende do solvente em que os corantes estão dissolvidos, ou seja, do tipo de moléculas do solvente com as quais as moléculas do corante estão associadas. Por exemplo, em uma planta viva, o principal pico de absorção da clorofila a na faixa vermelha está em 683 nm, deslocado em relação ao pico em 660 nm para essa clorofila dissolvida em éter etílico. Para a clorofila a, também foi encontrado um pico de absorção adicional em 672 nm. Presume-se, portanto, que isso seja resultado de pequenas alterações nas condições ambientais. Gráficos que mostram a dependência da absorção de luz em diferentes comprimentos de onda são chamados de espectros de absorção. O espectro de absorção é determinado usando um espectrofotômetro, ou seja, um instrumento equipado com uma fonte de luz que emite ondas nos comprimentos de onda desejados; um prisma ou uma grade de difração que serve para dispersar a luz; espelhos que refletem os comprimentos de onda selecionados e os direcionam para a amostra em teste; e um detector usado para detectar a quantidade de luz absorvida pela amostra em teste. A solução de clorofila irradiada Apresenta forte fluorescência, emitindo uma luz vermelha intensa. Como a maior parte da energia que causa a fluorescência em plantas vivas é utilizada na fotossíntese, elas não fluorescem visivelmente.
3. Síntese de clorofila.
A síntese de clorofila depende de diversos fatores, incluindo características genéticas, luz e a disponibilidade de certos elementos minerais. Em certas variedades de milho e girassol, algumas sementes em germinação produzem plântulas sem clorofila. Essa é uma característica recessiva inata. Esses brotos, chamados albinos, morrem quando as reservas nutricionais da semente se esgotam. No entanto, se essas plantas receberem açúcares como fonte de energia, podem continuar a crescer até quase a maturidade.
A luz é essencial para a síntese de clorofila em quase todas as angiospermas, embora coníferas, musgos, algumas samambaias e muitas algas também possam produzir esse pigmento no escuro. As angiospermas que crescem no escuro sintetizam pequenas quantidades de um pigmento verde semelhante à clorofila, mas seu teor é muito baixo para conferir a cor verde a essas plantas. Ao serem expostas à luz, esse pigmento desaparece, sendo substituído pela clorofila a, cuja quantidade aumenta com o tempo. Esse pigmento que desaparece é chamado de protoclorofila. Quimicamente, difere da clorofila a apenas pela ausência de dois átomos de hidrogênio na molécula. Além disso, absorve luz azul e vermelha de forma semelhante, apresentando máximos de absorção em comprimentos de onda de 450 nm e 650 nm. Esses comprimentos de onda da luz são os mais eficazes na conversão da protoclorofila em clorofila a. No entanto, a origem da clorofila b permanece inexplicada.
Ao que tudo indica, diversos elementos minerais participam diretamente da síntese da clorofila, visto que sua deficiência causa descoloração, ou clorose, nas plantas. Os anéis pirrólicos da clorofila são formados pela combinação prévia da glicina com o ácido succínico. Entre a reação desses dois compostos e a formação final do complexo clorofila, ocorre uma série de reações intermediárias catalisadas por enzimas específicas. A reação entre a glicina e o succinil-CoA depende da presença de ferro como coenzima, e a deficiência desse elemento causa clorose rápida em folhas jovens em desenvolvimento. Deficiências de manganês, potássio, zinco e cobre também causam clorose, mas o papel desses elementos ainda não é totalmente compreendido. Presume-se que eles ativem certas enzimas envolvidas na síntese da clorofila. A deficiência de manganês leva à destruição da estrutura lamelar interna dos cloroplastos e pode ser a razão para a inibição da síntese da clorofila que estabiliza as lamelas.
No entanto, a falta de nitrogênio e magnésio causa o amarelecimento das plantas. O papel desses elementos é compreensível, pois são os componentes estruturais básicos da clorofila. Isso não exclui suas funções mais indiretas; por exemplo, o magnésio provavelmente atua como coenzima em uma ou mais etapas da síntese da clorofila.
4. Estrutura e síntese de pigmentos carotenoides.
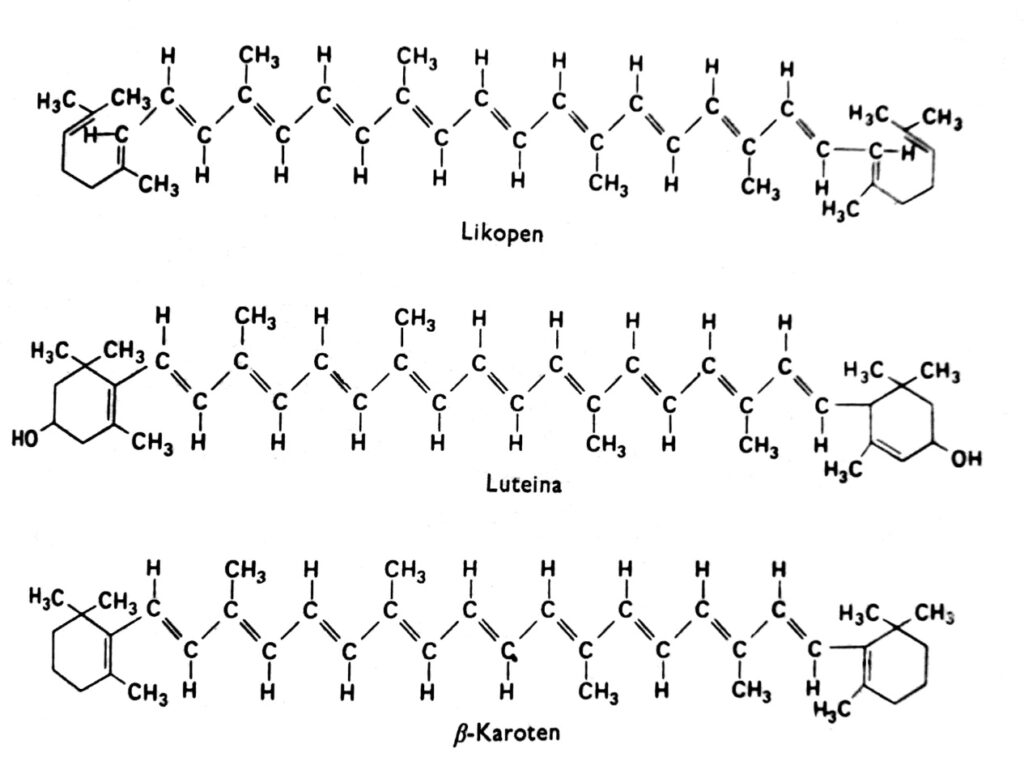
Os carotenoides são um grupo de pigmentos que, em plantas superiores, são geralmente vermelhos, laranjas, amarelos ou marrons e são encontrados nos cloroplastos em combinação com a clorofila. Eles também são encontrados, às vezes, em outros plastídeos, como os cromoplastos das raízes de cenoura e tomate, e nos cromoplastos de diversas flores amarelas. A cor amarela dos carotenoides é visível em angiospermas que crescem no escuro e não possuem clorofila (indicando que a luz não é necessária para seu acúmulo) e, no outono, nas folhas de muitas espécies, onde a clorofila desaparece. De fato, os carotenoides também podem exibir outras cores, como o verde em abacates, o rosa em camarões ou o preto intenso em cefalópodes. Esses pigmentos ocorrem em todos os principais grupos de plantas e animais, mas nossa discussão se concentrará principalmente naqueles encontrados nos cloroplastos em combinação com a clorofila. Os carotenoides podem ser divididos em dois grupos químicos: carotenos e xantofilas. Dentro de cada grupo, existem vários tipos distintos desses compostos, cujas moléculas são compostas por 40 átomos de carbono. Os carotenoides, cujo representante mais importante é o β-caroteno, são hidrocarbonetos (Fig. 2). As xantofilas, como a luteína e a zeaxantina, possuem átomos de oxigênio ligados aos seus anéis terminais. Tanto as xantofilas quanto os carotenos são sintetizados nas plantas a partir do acetato ativo, ou seja, acetil-CoA, via ácido mevalônico.
Figura 2. Fórmulas estruturais dos carotenoides.
Quase nenhum carotenoide se dissolve em água, mas eles se dissolvem em muitos solventes orgânicos, como acetona e éter. As xantofilas, por possuírem átomos de oxigênio adicionais, são mais hidrofílicas que os carotenos. Em solução, a maioria dos carotenoides absorve principalmente luz violeta e azul na faixa de 400-500 nm. Seus espectros de absorção dependem do grau de saturação das ligações duplas e, portanto, podem ser ligeiramente diferentes em cloroplastos intactos. Há evidências de que, em células intactas, o máximo de absorção de alguns carotenoides em 500 nm é deslocado para 450 nm após a extração desses pigmentos com um solvente orgânico.
5. Funções dos carotenoides.
Os carotenoides desempenham um papel importante na fotossíntese, provavelmente por dois motivos:
1. Eles desempenham uma função protetora, protegendo a clorofila dos efeitos nocivos da luz
e do oxigênio. Esse fenômeno destrutivo é chamado de foto-oxidação. É comum observar que uma solução alcoólica de clorofila se descolora sob a influência da luz, o que é acompanhado pelo consumo de oxigênio. Presumivelmente, uma reação semelhante ocorreria normalmente nas plantas se os carotenoides não cumprissem sua função protetora. A evidência mais convincente vem de observações de certos mutantes bacterianos. Como eles não contêm pigmentos carotenoides, a foto-oxidação ocorre muito facilmente neles. Além disso, sabe-se que mutantes albinos de girassol perderam a capacidade de sintetizar carotenoides, mas ainda produzem traços de clorofila. Nesse caso, o aumento da intensidade da luz desencadeia uma foto-oxidação adicional da clorofila, e a taxa desse processo é igual à sua síntese. O mecanismo pelo qual os carotenoides protegem a clorofila da oxidação permanece obscuro. No entanto, acredita-se que os carotenoides sejam foto-oxidados pela luz, o que poderia decompor a clorofila.
2. Outra função provável dos carotenoides, e especialmente das xantofilas, é o seu envolvimento na absorção de luz em comprimentos de onda ativos na fotossíntese. Há evidências indiscutíveis de que alguns carotenoides, como a fucoxantina (também chamada de fucoxantol), encontrada em algas pardas (por exemplo, fucus), participam ativamente da fotossíntese. Essa evidência foi obtida comparando-se o espectro de absorção da fucoxantina com o chamado espectro funcional, ou espectro da atividade fotossintética. O espectro funcional fotossintético é um gráfico que ilustra a eficiência da fotossíntese em função dos comprimentos de onda da luz que desencadeiam esse processo. Constatou-se que, em plantas verdes e plantas superiores, a luz absorvida por alguns carotenoides também é utilizada na fotossíntese. Nesses casos, os máximos do espectro funcional situam-se nas faixas de luz vermelha e azul, onde a clorofila absorve com maior intensidade. No entanto, a fotossíntese ainda ocorre usando luz azul-esverdeada, presumivelmente porque alguns carotenoides são ativos nessa faixa de comprimento de onda. A evidência mais convincente de que os carotenoides participam da fotossíntese é a sua capacidade de transferir energia de excitação diretamente para a clorofila a. Quando uma planta é irradiada com luz em comprimentos de onda absorvidos quase que exclusivamente pelos carotenoides, observa-se fluorescência vermelha da clorofila a. A hipótese é que a energia de excitação, às vezes liberada como fluorescência, seja tipicamente usada como força motriz para a fotossíntese.
6. Ficobilinas.
O último grupo de pigmentos fotossintéticos são as ficobilinas, encontradas em cianobactérias
e algas vermelhas. Esses pigmentos são classificados em dois grupos: vermelhos (ficoeritrinas) e azuis (ficocianinas). Assim como a clorofila, são compostos por quatro anéis pirrólicos. No entanto, as ficobilinas contêm um sistema de anéis pirrólicos aberto e não possuem magnésio nem um grupo fitol. Em células vivas, ocorrem associadas a proteínas e são solúveis em água. Ambos os tipos de pigmentos absorvem luz, utilizando-a para a fotossíntese. Em algas vermelhas, a maior eficiência fotossintética ocorre na faixa da luz verde, pois nessa faixa, a ficoeritrina, diferentemente da clorofila, absorve a luz com maior intensidade. Assim como os carotenoides, as ficobilinas transferem energia de excitação para a clorofila a, podendo causar sua fluorescência. Este modo de transferência de energia indica que as ficobilinas participam ativamente da fotossíntese ao capturar fótons, cuja energia é tipicamente utilizada pela clorofila a como força motriz para a fotossíntese. Pigmentos como carotenoides e ficobilinas, que transferem energia para um aceptor adequado, como a clorofila a, são chamados de pigmentos acessórios. O grupo de pigmentos acessórios também inclui as clorofilas b, c e d, porque elas absorvem energia luminosa, que por sua vez pode ser transferida para a clorofila a.
Literatura:
-Franck B. Salibury, Cleon Ross "Fisiologia Vegetal" PWRiL 1975.
-Otis F. Daniel G. Curtis Clark "Introdução à Fisiologia Vegetal" PWRiL 1958.
-Zurzycki J. Michniewicz M. "Fisiologia Vegetal" PWRiL 1979.
-Richter G. "Processos metabólicos em plantas" PWN 1975.