1. Bevezetés.
A fotoszintézis minden algában és magasabb rendű növényben klorofill - a jelenlétében megy végbe ; kivételt képeznek a fotoszintetizáló baktériumok, amelyek a klorofill- a- tól kissé eltérő szerkezetű pigmenteket tartalmaznak . A klorofill más fajtái is előfordulnak a növényekben.
A magasabb rendű növények és a zöldalgák klorofill a-t és b-t, míg a cianobaktériumok és egyes vörösalgák csak klorofill a-t. Más vörösalgák klorofill a-t és kis mennyiségű klorofill d-t, míg a barnaalgák és a kovamoszatok klorofill a-tés klorofill c-t.
Ezenkívül más pigmentek is részt vesznek a fotoszintézisben, például a karotinoidok, valamint a vörös algákban és cianobaktériumokban található fikobilin pigmentek (fikoeritrin és fikocianin). Az 1. táblázat felsorolja a különböző fotoszintetikus organizmusokban található pigmenteket, beleértve azok színét és elsődleges abszorpciós maximumát. Az 1. ábra néhány fotoszintetikus pigment alapvető kémiai szerkezetét mutatja.
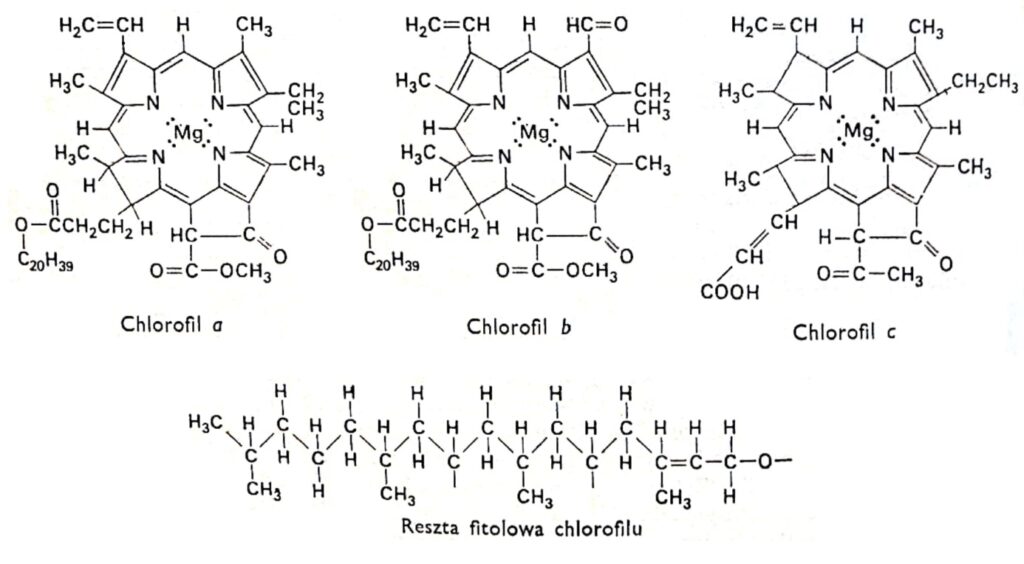
1. ábra. A klorofillok szerkezeti képletei.
1. táblázat. A fotoszintetikus pigmentek tulajdonságai.
Festék | Szín | Fő abszorpciós csúcsok a látható tartományban (nm) |
| Klorofill a | kékeszöld | 430 (kék-ibolya) 660 (piros) |
| Klorofill b | zöld | 455 (kék) 640 (narancssárga-piros) |
| fikocianinok | kék | 560-660 (sárgától narancsvörösig) |
| Fikoeritrinek | piros | 550-570 (zöldből sárgába) |
| Karotinoidok | Sárga, narancsvörös | 430-490 (lilától kékeszöldig) |
2. A klorofill szerkezete.
A klorofill minden változatának azonos porfirin szerkezete van, amely négy pirrolgyűrűből áll, amelyek nitrogénatomokon keresztül kapcsolódnak a magnéziumhoz. Továbbá a klorofill molekula tartalmaz egy ötödik gyűrűt, amely teljes egészében szénatomokból áll, és a legtöbb klorofillre jellemző hosszú szénláncú alkohol fitolhoz kapcsolódik, a klorofill c kivételével . A klorofillok a pirrolgyűrűkhöz kapcsolódó oldalláncok szerkezetében különböznek. A klorofill a és b közötti különbség az, hogy a klorofill a harmadik gyűrűje metilcsoportot tartalmaz, míg a klorofill b -ben aldehidcsoportot. Az aldehidcsoport hidrofilebbé teszi a klorofill b-t (92%-os metil-alkoholban oldódik), mint a klorofill a-t (jobban oldódik éterben). A növények túlnyomó többsége kétszer-háromszor több klorofill a-t tartalmaz, mint klorofill b-t . A klorofill molekula szerkezetét a század elején a német kutatók, R. Wilstatter és A. Stoll, valamint a híres szerves kémikus, Hans Fischer határozták meg.
Az a és b klorofill elsősorban kékes-ibolya és vörös fényt nyel el, és csak kissé a zöld fényt. A klorofill a esetében a maximális abszorpció az ibolya tartományban történik, azaz rövidebb hullámhosszakon, mint a b klorofill esetében ; míg a vörös tartományban a maximum a hosszabb hullámhosszak felé tolódik el. „Az abszorpciós maximum helye attól az oldószertől függ, amelyben a festékanyagok feloldódnak, azaz az oldószermolekulák típusától, amelyekkel a festékmolekulák kapcsolatban állnak. Például egy élő növényben a klorofill a fő abszorpciós csúcsa a vörös tartományban 683 nm-nél van, amely eltolódik az etil-éterben oldott klorofill 660 nm-es csúcsához képest. A klorofill a esetében egy további abszorpciós csúcsot is találtak 672 nm-nél. Ezért feltételezzük, hogy ez a kissé megváltozott környezeti feltételek eredménye. A fényelnyelés különböző hullámhosszaktól való függését bemutató grafikonokat abszorpciós spektrumoknak nevezzük. Az abszorpciós spektrumot spektrofotométerrel, azaz egy olyan műszerrel határozzuk meg, amely a kívánt hullámhosszúságú hullámokat kibocsátó fényforrással van felszerelve; egy prizmával vagy diffrakciós ráccsal, amely a fényt szétszórja; tükrökkel, amelyek a kiválasztott hullámhosszakat visszaverik és a vizsgált mintára irányítják; és egy detektorral, amelyet a vizsgált minta által elnyelt fény mennyiségének mérésére használnak. A besugárzott klorofilloldat erősen fluoreszkál, intenzív vörös fényt bocsátva ki. Mivel az élő növényekben a fluoreszcenciát okozó energia nagy része…” fotoszintézisben használják, láthatóan nem fluoreszkálnak.
3. Klorofill szintézise.
A klorofill szintézise számos tényezőtől függ, beleértve a genetikai jellemzőket, a fényt és bizonyos ásványi elemek elérhetőségét. Bizonyos kukorica- és napraforgófajtákban egyes csírázó magok klorofillhiányos palántákat hoznak. Ez egy veleszületett recesszív tulajdonság. Ezek a hajtások, az úgynevezett albínók, elpusztulnak, amikor a mag tápanyagtartalékai kimerülnek. Ha azonban ezeket a növényeket cukrokkal látják el energiaforrásként, akkor majdnem az érettségig képesek növekedni.
A fény elengedhetetlen a klorofill szintéziséhez szinte minden zárvatermőben, bár a tűlevelűek, mohák, egyes páfrányok és sok alga is képes ezt a pigmentet sötétben termelni. A sötétben növő zárvatermők kis mennyiségben szintetizálnak egy, a klorofillhoz hasonló zöld pigmentet, de ennek tartalma túl alacsony ahhoz, hogy zöld színt kölcsönözzön ezeknek a növényeknek. Fény hatására ez a pigment eltűnik, helyét klorofill a veszi át , amelynek mennyisége idővel növekszik. Ezt az eltűnő pigmentet protoklorofillnek nevezik. Kémiailag csak abban különbözik a klorofill a-tól, hogy nincs két hidrogénatom a molekulában. Továbbá hasonlóan nyeli el a kék és a vörös fényt, abszorpciós maximumot mutatva 450 nm és 650 nm hullámhosszon. Ezek a fényhullámhosszak a leghatékonyabbak a protoklorofill klorofill a -vá alakításában. A klorofill b eredete azonban továbbra sem tisztázott.
Úgy tűnik, hogy számos ásványi elem közvetlenül részt vesz a klorofill szintézisében, mivel hiányuk elszíneződést, vagyis klorózist okoz a növényekben. A klorofill-pirrolgyűrűk a glicin és a borostyánkősav előzetes kombinációjából képződnek. E két vegyület reakciója és a klorofill komplex vegyület végső képződése között egy sor köztes reakció játszódik le, amelyeket specifikus enzimek katalizálnak. A glicin és a szukcinil-CoA közötti reakció a vas koenzim jelenlététől függ, és ennek az elemnek a hiánya gyors klorózist okoz a fiatal, fejlődő levelekben. A mangán, a kálium, a cink és a réz hiánya szintén klorózist okoz, de ezen elemek szerepe még nem ismert. Feltehetően aktiválják a klorofill szintézisében részt vevő bizonyos enzimeket. A mangánhiány a kloroplasztok belső lemezes szerkezetének pusztulásához vezet, és oka lehet a lamellákat stabilizáló klorofill szintézisének gátlásának.
A nitrogén és a magnézium hiánya azonban a növények sárgulását okozza. Ezen elemek szerepe érthető, mivel ezek a klorofill alapvető szerkezeti alkotóelemei. Ez nem zárja ki közvetettebb funkcióikat, például a magnézium valószínűleg koenzimként működik a klorofillszintézis egy vagy több lépésében.
4. Karotinoid pigmentek szerkezete és szintézise .
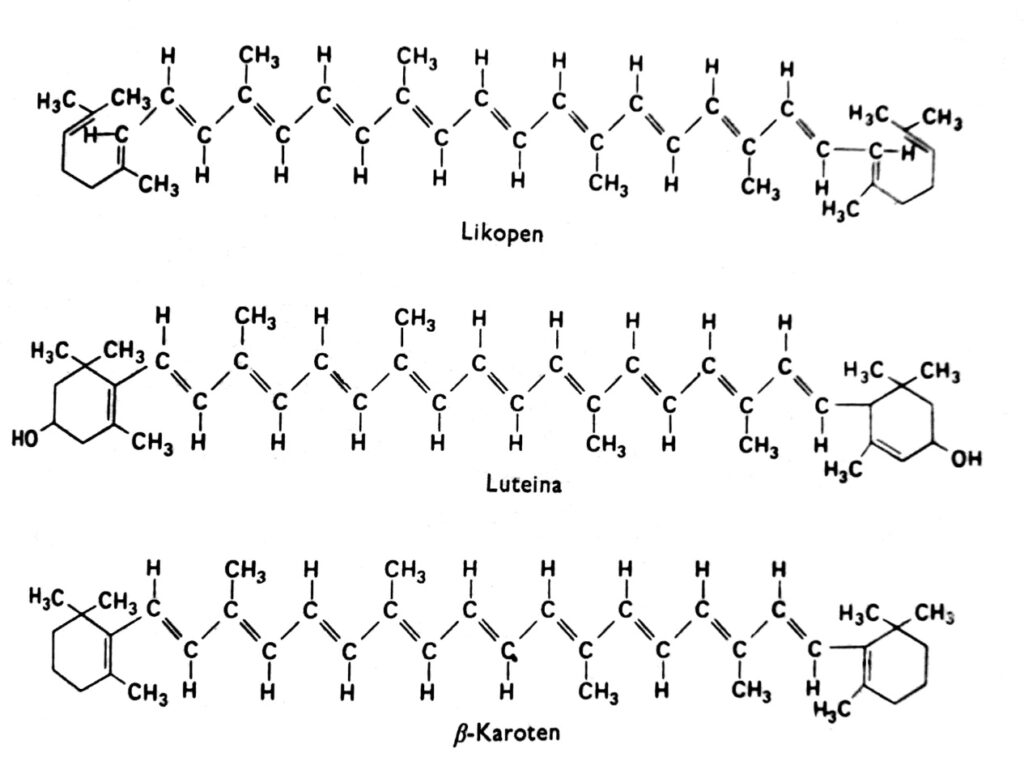
A karotinoidok olyan pigmentek csoportja, amelyek a magasabb rendű növényekben általában vörös, narancssárga, sárga vagy barna színűek, és klorofillal kombinálva találhatók a kloroplasztokban. Néha más plasztidokban is megtalálhatók, például a sárgarépa és a paradicsom gyökereinek kromoplasztjaiban, valamint számos sárga virág kromoplasztjaiban. A karotinoidok sárga színe látható a sötétben növő és klorofillmentes zárvatermőkben (ami azt jelzi, hogy a fény nem szükséges a felhalmozódáshoz), valamint ősszel számos faj levelében, ahol a klorofill eltűnik. Valójában a karotinoidok más színeket is mutathatnak, például zöldet az avokádóban, rózsaszínt a garnélában, vagy majdnem tintafeketét a lábasfejűekben. Ezek a pigmentek a növények és állatok minden főbb csoportjában előfordulnak, de a mi megbeszélésünk elsősorban azokra összpontosít, amelyek kloroplasztokban találhatók klorofillal kombinálva. A karotinoidok két kémiai csoportra oszthatók: karotinokra és xantofillekre. Mindegyik csoporton belül ezeknek a vegyületeknek több különböző típusa létezik, amelyek molekulái 40 szénatomból állnak. A karotinoidok, melyek legfontosabb képviselője a β-karotin, szénhidrogének (2. ábra). A xantofillok, mint például a lutein és a zeaxantin, oxigénatomokat tartalmaznak a terminális gyűrűikhez kapcsolódva. Mind a xantofilleket, mind a karotinokat a növények aktív acetátból, azaz acetil-CoA-ból szintetizálják mevalonsav útján.
2. ábra. A karotinoidok szerkezeti képletei.
A karotinoidok szinte egyike sem oldódik vízben, viszont számos szerves oldószerben, például acetonban és éterben oldódnak. A további oxigénatomokat tartalmazó xantofillek hidrofilebbek, mint a karotinok. Oldatban a legtöbb karotinoid elsősorban az ibolya és kék fényt nyeli el a 400-500 nm-es tartományban. Abszorpciós spektrumuk a kettős kötés telítettségének mértékétől függ, ezért kissé eltérhet az ép kloroplasztokban. Bizonyítékok vannak arra, hogy ép sejtekben egyes karotinoidok abszorpciós maximuma 500 nm-en 450 nm-re tolódik el ezen pigmentek szerves oldószeres extrakciója után.
5. A karotinoidok funkciói.
A karotinoidok fontos szerepet játszanak a fotoszintézisben, valószínűleg két okból:
1. Védőfunkciót látnak el, megvédik a klorofillt a fény
és az oxigén káros hatásaitól. Ezt a romboló jelenséget fotooxidációnak nevezik. Gyakran megfigyelhető, hogy a klorofill alkoholos oldata fény hatására elszíneződik, amit oxigénfogyasztás kísér. Feltehetően hasonló reakció játszódna le normális esetben a növényekben is, ha a karotinoidok nem töltenék be védőfunkciójukat. A legmeggyőzőbb bizonyíték bizonyos baktériummutánsok megfigyeléseiből származik. Mivel nem tartalmaznak karotinoid pigmenteket, a fotooxidáció nagyon könnyen végbemegy bennük. Ismertek továbbá olyan albínó napraforgómutánsok, amelyek elvesztették a karotinoidok szintetizálásának képességét, de nyomokban még termelnek klorofillt. Ebben az esetben a növekvő fényintenzitás további klorofill fotooxidációját váltja ki, és ennek a folyamatnak a sebessége megegyezik a szintézisével. A karotinoidok klorofill oxidációtól való védelmének mechanizmusa továbbra sem tisztázott. Úgy vélik azonban, hogy a karotinoidokat fény fotooxidálja, ami lebonthatja a klorofillt.
2. A karotinoidok, és különösen a xantofillek egy másik valószínűsíthető funkciója a fotoszintézisben aktív hullámhosszú fényelnyelésében való részvételük. Vitathatatlan bizonyítékok vannak arra, hogy egyes karotinoidok, mint például a barna algákban (pl. fucusban) található fukoxantin (más néven fukoxantol), aktívan részt vesznek a fotoszintézisben. Ezt a bizonyítékot a fukoxantin abszorpciós spektrumának az úgynevezett funkcionális spektrummal, vagyis a fotoszintetikus aktivitás spektrumával való összehasonlításával szerezték. A fotoszintetikus funkcionális spektrum egy grafikon, amely a fotoszintézis hatékonyságát szemlélteti a folyamatot kiváltó fény hullámhosszainak függvényében. Megállapították, hogy zöld növényekben és magasabb rendű növényekben egyes karotinoidok által elnyelt fényt a fotoszintézis is felhasználja. Ilyen esetekben a funkcionális spektrum maximumai a vörös és kék fénytartományba esnek, ahol a klorofill a legerősebben nyeli el. A fotoszintézis azonban továbbra is kékeszöld fény segítségével történik, feltehetően azért, mert egyes karotinoidok ebben a tartományban aktívak. A karotinoidok fotoszintézisben való részvételének legmeggyőzőbb bizonyítéka az a képességük, hogy gerjesztett energiát közvetlenül a klorofill a-nak adnak át. Amikor egy növényt olyan hullámhosszú fénnyel besugároznak, amelyet szinte kizárólag karotinoidok abszorbeálnak, a klorofill a vörös fluoreszcenciáját figyelik meg. A feltételezés szerint a gerjesztési energia, amely néha fluoreszcencia formájában szabadul fel, jellemzően a fotoszintézis hajtóerejeként szolgál.
6. Fikobilinek.
A fotoszintetikus pigmentek utolsó csoportját a fikobilinek alkotják, amelyek cianobaktériumokban
és vörös algákban találhatók. Ezeket a pigmenteket két csoportba sorolják: vörös (fikoeritrinek) és kék (fikocianinok). A klorofillhoz hasonlóan négy pirrolgyűrűből állnak. A fikobilinek azonban nyitott pirrolgyűrűs rendszert tartalmaznak, és sem magnéziumot, sem fitolrészt nem tartalmaznak. Élő sejtekben fehérjékkel együtt fordulnak elő, és vízben oldódnak. Mindkét típusú pigment elnyeli a fényt, és fotoszintézishez használja azt. A vörös algákban a legnagyobb fotoszintetikus hatékonyság a zöld fénytartományban fordul elő, mivel ebben a tartományban a fikoeritrin, a klorofillal ellentétben, a legintenzívebben nyeli el a fényt. A karotinoidokhoz hasonlóan a fikobilinek gerjesztett energiát adnak át a klorofill a-nak, és fluoreszkálást okozhatnak. Ez az energiaátadási mód azt jelzi, hogy a fikobilinek aktívan részt vesznek a fotoszintézisben a fotonok befogásával, amelyek energiáját jellemzően a klorofill a a fotoszintézis hajtóerejeként használja fel. Az olyan pigmenteket, mint a karotinoidok és a fikobilinek, amelyek energiát adnak át egy megfelelő akceptornak, pl. a klorofill a-, járulékos pigmenteknek nevezzük. A járulékos pigmentek csoportjába tartoznak a b, c és d, mivel ezek elnyelik a fényenergiát, amely viszont átvihető a klorofill a.
Irodalom:
-Franck B. Salibury, Cleon Ross „Növényélettan” PWRiL 1975.
-Otis F. Daniel G. Curtis Clark „Bevezetés a növényélettanba” PWRiL 1958.
-Zurzycki J. Michniewicz M. "Növényfiziológia" PWRiL 1979.
-Richter G. "Anyaganyalatok növényekben" PWN 1975.