1. Introduction.
La photosynthèse chez toutes les algues et les plantes supérieures se déroule en présence de chlorophylle a; les bactéries photosynthétiques font exception, car elles contiennent des pigments dont la structure est légèrement différente de celle de la chlorophylle a. D'autres types de chlorophylle sont également présents chez les plantes.
Les plantes supérieures et les algues vertes contiennent de la chlorophylle a et b, tandis que les cyanobactéries et certaines algues rouges ne contiennent que de la chlorophylle a. D'autres espèces d'algues rouges contiennent de la chlorophylle a et une petite quantité de chlorophylle d, et les algues brunes et les diatomées contiennent à la fois de la chlorophylle aet de la chlorophylle c.
D'autres pigments participent également à la photosynthèse, comme les caroténoïdes, et chez les algues rouges et les cyanobactéries, les phycobilines (phycoérythrine et phycocyanine). Le tableau 1 répertorie les pigments présents chez divers organismes photosynthétiques, en précisant leur couleur et leur longueur d'onde d'absorption maximale. La figure 1 illustre la structure chimique de base de certains pigments photosynthétiques.
Fig. 1. Formules structurales des chlorophylles.
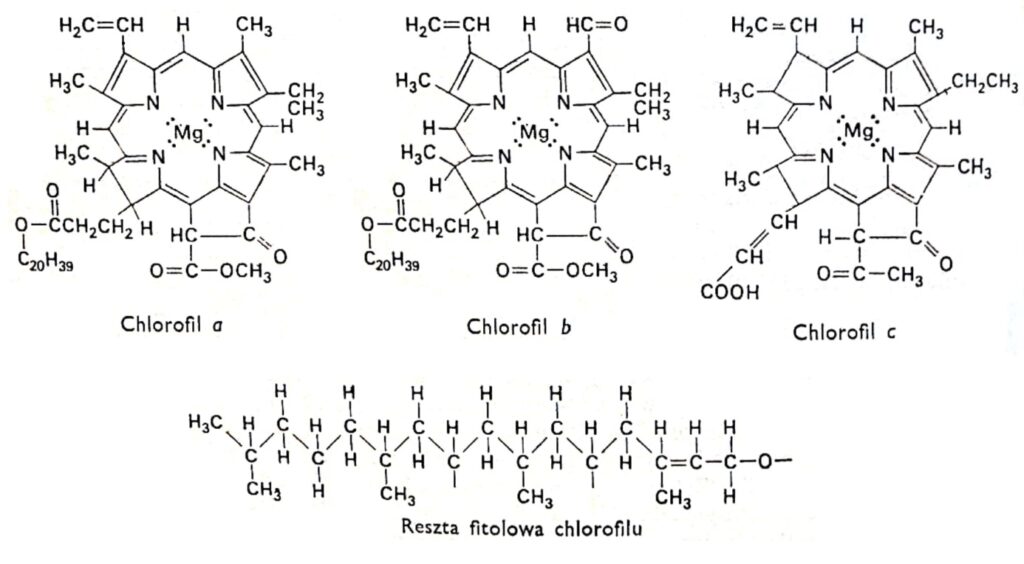
Tableau 1. Propriétés des pigments photosynthétiques.
Colorant | Couleur | Principaux pics d'absorption dans le domaine visible (nm) |
| Chlorophylle a | bleu-vert | 430 (bleu-violet) 660 (rouge) |
| Chlorophylle b | vert | 455 (bleu) 640 (orange-rouge) |
| phycocyanines | bleu | 560-660 (jaune à rouge orangé) |
| Phycoérythrines | rouge | 550-570 (vert à jaune) |
| Caroténoïdes | Jaune, rouge orangé | 430-490 (violet à bleu-vert) |
2. La structure de la chlorophylle.
Toutes les variétés de chlorophylle possèdent la même structure porphyrinique, composée de quatre cycles pyrrole liés au magnésium par des atomes d'azote. De plus, la molécule de chlorophylle contient un cinquième cycle, entièrement composé d'atomes de carbone et lié au phytol, un alcool à longue chaîne, caractéristique de la plupart des chlorophylles, à l'exception de la chlorophylle c. Les chlorophylles diffèrent par la structure des chaînes latérales fixées aux cycles pyrrole. La différence entre la chlorophylle a et b réside dans le fait que le troisième cycle de la chlorophylle a contient un groupe méthyle, tandis que celui de la chlorophylle b contient un groupe aldéhyde. Ce groupe aldéhyde rend la chlorophylle b plus hydrophile (soluble dans le méthanol à 92 %) que la chlorophylle a (plus soluble dans l'éther). La grande majorité des plantes contiennent deux à trois fois plus de chlorophylle a que de chlorophylle b. La structure de la molécule de chlorophylle a été déterminée au début du XXe siècle par les chercheurs allemands R. Wilstatter et A. Stoll, ainsi que par le célèbre chimiste organicien Hans Fischer.
Les chlorophylles a et b absorbent principalement la lumière bleu-violet et rouge, et
très peu la lumière verte. Pour la chlorophylle a, l'absorption maximale se situe dans le domaine violet, c'est-à-dire à des longueurs d'onde plus courtes que pour la chlorophylle b ; tandis que dans le domaine rouge, le maximum est décalé vers les longueurs d'onde plus longues. La position du maximum d'absorption dépend du solvant dans lequel les colorants sont dissous, c'est-à-dire du type de molécules de solvant auxquelles les molécules de colorant sont associées. Par exemple, dans une plante vivante, le pic d'absorption principal de la chlorophylle a dans le rouge se situe à 683 nm, soit un décalage par rapport au pic à 660 nm de cette même chlorophylle dissoute dans l'éther éthylique. Pour la chlorophylle a, un pic d'absorption supplémentaire a également été observé à 672 nm. On suppose donc que ce décalage résulte de légères modifications des conditions environnementales. Les graphiques illustrant la dépendance de l'absorption de la lumière à différentes longueurs d'onde sont appelés spectres d'absorption. Le spectre d'absorption est déterminé à l'aide d'un spectrophotomètre, instrument équipé d'une source lumineuse émettant des ondes aux longueurs d'onde souhaitées ; d'un prisme ou d'un réseau de diffraction servant à disperser la lumière ; de miroirs réfléchissant certaines longueurs d'onde et les dirigeant vers l'échantillon analysé ; et d'un détecteur permettant de mesurer la quantité de lumière absorbée par l'échantillon. La solution de chlorophylle irradiée devient fluorescente. Elles émettent fortement une lumière rouge intense. Comme la majeure partie de l'énergie responsable de la fluorescence chez les plantes vivantes est utilisée pour la photosynthèse, elles ne présentent pas de fluorescence visible.
3. Synthèse de la chlorophylle.
La synthèse de la chlorophylle dépend de plusieurs facteurs, notamment des caractéristiques génétiques, de la lumière et de la disponibilité de certains éléments minéraux. Chez certaines variétés de maïs et de tournesol, certaines graines en germination produisent des plantules dépourvues de chlorophylle. Il s'agit d'un caractère récessif inné. Ces pousses, dites albinos, meurent lorsque les réserves nutritives de la graine sont épuisées. Cependant, si ces plantes reçoivent des sucres comme source d'énergie, elles peuvent continuer à croître jusqu'à une quasi-maturité.
La lumière est essentielle à la synthèse de la chlorophylle chez presque toutes les angiospermes, bien que les conifères, les mousses, certaines fougères et de nombreuses algues puissent également produire ce pigment à l'obscurité. Les angiospermes qui poussent dans l'obscurité synthétisent de faibles quantités d'un pigment vert semblable à la chlorophylle, mais sa concentration est insuffisante pour conférer une couleur verte à ces plantes. Sous l'effet de la lumière, ce pigment disparaît et est remplacé par la chlorophylle a, dont la quantité augmente avec le temps. Ce pigment qui disparaît est appelé protochlorophylle. Chimiquement, il ne diffère de la chlorophylle a que par l'absence de deux atomes d'hydrogène dans sa molécule. De plus, il absorbe la lumière bleue et rouge de manière similaire, présentant des maxima d'absorption aux longueurs d'onde de 450 nm et 650 nm. Ces longueurs d'onde sont les plus efficaces pour la conversion de la protochlorophylle en chlorophylle a. Cependant, l'origine de la chlorophylle b demeure inexpliquée.
Il apparaît que plusieurs éléments minéraux participent directement à la synthèse de la chlorophylle, leur carence provoquant une décoloration, ou chlorose, chez les plantes. Les cycles pyrrole de la chlorophylle sont formés par la combinaison préalable de la glycine et de l'acide succinique. Entre la réaction de ces deux composés et la formation finale de la chlorophylle, une série de réactions intermédiaires catalysées par des enzymes spécifiques se produisent. La réaction entre la glycine et le succinyl-CoA dépend de la présence de fer comme coenzyme, et une carence en cet élément provoque une chlorose rapide des jeunes feuilles en développement. Les carences en manganèse, potassium, zinc et cuivre provoquent également une chlorose, mais le rôle de ces éléments reste encore inconnu. Ils activent probablement certaines enzymes impliquées dans la synthèse de la chlorophylle. Une carence en manganèse entraîne la destruction de la structure lamellaire interne des chloroplastes et pourrait être à l'origine de l'inhibition de la synthèse de la chlorophylle, stabilisant ainsi les lamelles.
Cependant, une carence en azote et en magnésium provoque le jaunissement des plantes. Le rôle de ces éléments est compréhensible, puisqu'ils constituent les éléments structuraux de base de la chlorophylle. Cela n'exclut pas leurs fonctions plus indirectes ; par exemple, le magnésium agit probablement comme coenzyme dans une ou plusieurs étapes de la synthèse de la chlorophylle.
4. Structure et synthèse des pigments caroténoïdes.
Les caroténoïdes sont un groupe de pigments qui, chez les plantes supérieures, sont généralement rouges, oranges, jaunes ou bruns et se trouvent dans les chloroplastes en association avec la chlorophylle. On les trouve aussi parfois dans d'autres plastes, comme les chromoplastes des racines de carotte et de tomate, et dans les chromoplastes de nombreuses fleurs jaunes. La couleur jaune des caroténoïdes est visible chez les angiospermes qui poussent à l'obscurité et en l'absence de chlorophylle (ce qui indique que la lumière n'est pas nécessaire à leur accumulation) et en automne dans les feuilles de nombreuses espèces, lorsque la chlorophylle disparaît. En effet, les caroténoïdes peuvent aussi présenter d'autres couleurs, comme le vert chez les avocats, le rose chez les crevettes ou un noir presque d'encre chez les céphalopodes. Ces pigments sont présents dans tous les grands groupes de plantes et d'animaux, mais nous nous concentrerons ici principalement sur ceux qui se trouvent dans les chloroplastes en association avec la chlorophylle. Les caroténoïdes se divisent en deux groupes chimiques : les carotènes et les xanthophylles. Au sein de chaque groupe, on distingue plusieurs types de ces composés, dont les molécules sont composées de 40 atomes de carbone. Les caroténoïdes, dont le représentant le plus important est le β-carotène, sont des hydrocarbures (Fig. 2). Les xanthophylles, comme la lutéine et la zéaxanthine, possèdent des atomes d'oxygène sur leurs cycles terminaux. Les xanthophylles et les carotènes sont synthétisés chez les plantes à partir d'acétate actif, c'est-à-dire l'acétyl-CoA, via l'acide mévalonique.
Fig. 2. Formules structurales des caroténoïdes.
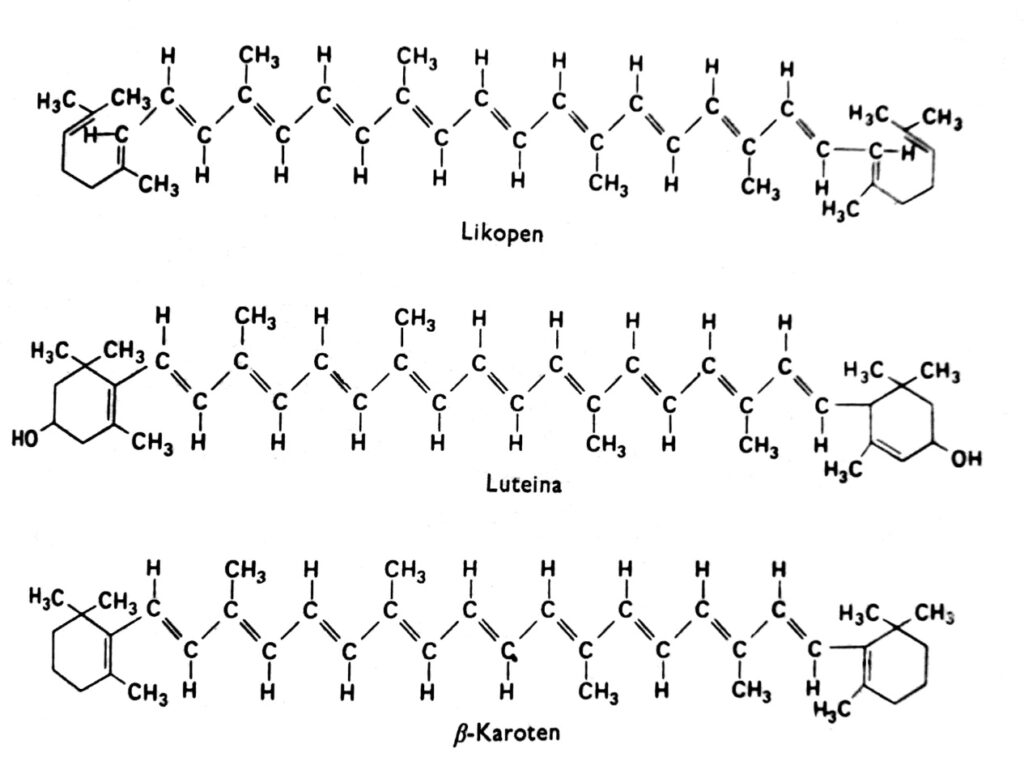
Presque aucun caroténoïde ne se dissout dans l'eau, mais ils sont solubles dans de nombreux solvants organiques, tels que l'acétone et l'éther. Les xanthophylles, possédant des atomes d'oxygène supplémentaires, sont plus hydrophiles que les carotènes. En solution, la plupart des caroténoïdes absorbent principalement la lumière violette et bleue dans la gamme 400-500 nm. Leurs spectres d'absorption dépendent du degré de saturation des doubles liaisons et peuvent donc présenter de légères différences dans les chloroplastes intacts. Il a été démontré que, dans les cellules intactes, le maximum d'absorption de certains caroténoïdes à 500 nm est déplacé à 450 nm après extraction de ces pigments par un solvant organique.
5. Fonctions des caroténoïdes.
Les caroténoïdes jouent un rôle important dans la photosynthèse, probablement pour deux raisons :
1. Ils exercent une fonction protectrice, préservant la chlorophylle des effets néfastes de la lumière
et de l'oxygène. Ce phénomène destructeur est appelé photo-oxydation. On observe souvent qu'une solution alcoolique de chlorophylle se décolore sous l'effet de la lumière, ce qui s'accompagne d'une consommation d'oxygène. Vraisemblablement, une réaction similaire se produirait normalement chez les plantes si les caroténoïdes n'assuraient pas leur fonction protectrice. La preuve la plus convaincante provient des observations réalisées sur certains mutants bactériens. Dépourvus de pigments caroténoïdes, ces mutants sont très sensibles à la photo-oxydation. De plus, on connaît des mutants albinos de tournesol qui ont perdu la capacité de synthétiser les caroténoïdes mais produisent encore des traces de chlorophylle. Dans ce cas, l'augmentation de l'intensité lumineuse déclenche une photo-oxydation supplémentaire de la chlorophylle, et la vitesse de ce processus est égale à celle de sa synthèse. Le mécanisme par lequel les caroténoïdes protègent la chlorophylle de l'oxydation reste encore mal compris. Cependant, on pense que les caroténoïdes sont photo-oxydés par la lumière, ce qui pourrait décomposer la chlorophylle.
2. Une autre fonction probable des caroténoïdes, et notamment des xanthophylles, est leur implication dans l'absorption de la lumière aux longueurs d'onde actives pour la photosynthèse. Il est incontestable que certains caroténoïdes, comme la fucoxanthine (ou fucoxanthol), présente dans les algues brunes (par exemple, le fucus), participent activement à la photosynthèse. Cette preuve a été obtenue en comparant le spectre d'absorption de la fucoxanthine au spectre fonctionnel, ou spectre d'activité photosynthétique. Ce dernier est un graphique illustrant l'efficacité de la photosynthèse en fonction des longueurs d'onde de la lumière qui déclenchent ce processus. On a constaté que chez les plantes vertes et les plantes supérieures, la lumière absorbée par certains caroténoïdes est également utilisée pour la photosynthèse. Dans ce cas, les maxima du spectre fonctionnel se situent dans les gammes du rouge et du bleu, où la chlorophylle absorbe le plus fortement. Cependant, la photosynthèse a également lieu en utilisant la lumière bleu-vert, probablement parce que certains caroténoïdes sont actifs dans cette gamme. L'argument le plus convaincant en faveur de la participation des caroténoïdes à la photosynthèse réside dans leur capacité à transférer directement l'énergie d'excitation à la chlorophylle a. Lorsqu'une plante est irradiée par une lumière dont les longueurs d'onde sont absorbées presque exclusivement par les caroténoïdes, on observe une fluorescence rouge de la chlorophylle a. L'hypothèse est que l'énergie d'excitation, parfois libérée sous forme de fluorescence, est généralement utilisée comme force motrice de la photosynthèse.
6. Phycobilines.
Le dernier groupe de pigments photosynthétiques est celui des phycobilines, présentes chez les cyanobactéries
et les algues rouges. Ces pigments se divisent en deux groupes : les phycoérythrines rouges et les phycocyanines bleues. Comme la chlorophylle, elles sont composées de quatre cycles pyrrole. Cependant, les phycobilines possèdent un système cyclique pyrrole ouvert et ne contiennent ni magnésium ni groupement phytol. Dans les cellules vivantes, elles sont associées à des protéines et sont solubles dans l’eau. Ces deux types de pigments absorbent la lumière et l’utilisent pour la photosynthèse. Chez les algues rouges, l’efficacité photosynthétique est maximale dans le spectre de la lumière verte, car c’est dans ce spectre que la phycoérythrine, contrairement à la chlorophylle, absorbe la lumière le plus intensément. À l’instar des caroténoïdes, les phycobilines transfèrent l’énergie d’excitation à la chlorophylle a et peuvent induire sa fluorescence. Ce mode de transfert d’énergie indique que les phycobilines participent activement à la photosynthèse en capturant les photons, dont l’énergie est généralement utilisée par la chlorophylle a comme force motrice de la photosynthèse. Les pigments tels que les caroténoïdes et les phycobilines, qui transfèrent de l'énergie à un accepteur approprié, par exemple la chlorophylle a, sont appelés pigments accessoires. Ce groupe comprend également les chlorophylles b, c et d, car elles absorbent l'énergie lumineuse, qui peut ensuite être transférée à la chlorophylle a.
Littérature:
-Franck B. Salibury, Cleon Ross « Physiologie végétale » PWRiL 1975.
-Otis F. Daniel G. Curtis Clark « Introduction à la physiologie végétale » PWRiL 1958.
-Zurzycki J. Michniewicz M. "Physiologie végétale" PWRiL 1979.
-Richter G. « Processus métaboliques chez les plantes » PWN 1975.