1. Introducción.
La fotosíntesis en todas las algas y plantas superiores se produce en presencia de clorofila a; la excepción son las bacterias fotosintéticas, que contienen pigmentos con una estructura ligeramente diferente a la de la clorofila a. En las plantas también se encuentran otras variedades de clorofila.
Las plantas superiores y las algas verdes contienen clorofila a y b, mientras que las cianobacterias y algunas algas rojas contienen solo clorofila a. Otras especies de algas rojas contienen clorofila a y una pequeña cantidad de clorofila d, y las algas pardas y las diatomeas tienen tanto clorofila acomo clorofila c.
Además, otros pigmentos participan en la fotosíntesis, como los carotenoides, y en las algas rojas y cianobacterias, los pigmentos ficobilinos (ficoeritrina y ficocianina). La Tabla 1 enumera los pigmentos presentes en diversos organismos fotosintéticos, incluyendo su color y máximo de absorción primaria. La Figura 1 muestra las estructuras químicas básicas de algunos pigmentos fotosintéticos.
Figura 1. Fórmulas estructurales de las clorofilas.
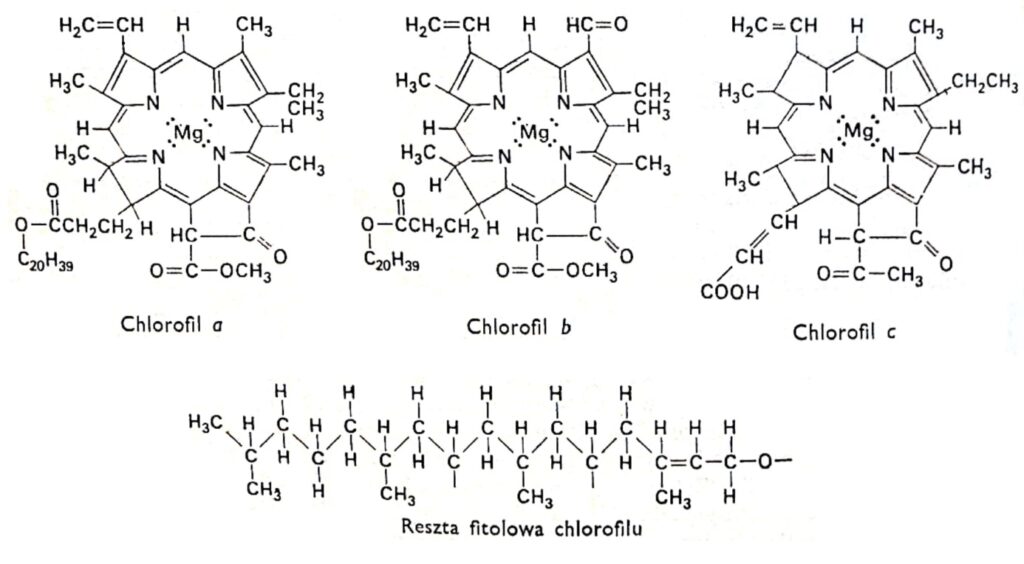
Tabla 1. Propiedades de los pigmentos fotosintéticos.
Teñir | Color | Principales picos de absorción en el rango visible (nm) |
| Clorofila a | azul verdoso | 430 (azul-violeta) 660 (rojo) |
| Clorofila b | verde | 455 (azul) 640 (rojo anaranjado) |
| Ficocianinas | azul | 560-660 (amarillo a rojo anaranjado) |
| Ficoeritrinas | rojo | 550-570 (verde a amarillo) |
| Carotenoides | Amarillo, rojo anaranjado | 430-490 (púrpura a azul verdoso) |
2. La estructura de la clorofila.
Todas las variedades de clorofila tienen la misma estructura de porfirina, compuesta por cuatro anillos de pirrol unidos al magnesio mediante átomos de nitrógeno. Además, la molécula de clorofila contiene un quinto anillo compuesto enteramente de átomos de carbono y unido al fitol, un alcohol de cadena larga, típico de la mayoría de las clorofilas, con la excepción de la clorofila c. Las clorofilas difieren en la estructura de las cadenas laterales unidas a los anillos de pirrol. La diferencia entre la clorofila a y la b es que el tercer anillo de la clorofila a contiene un grupo metilo, mientras que en la clorofila b contiene un grupo aldehído. El grupo aldehído hace que la clorofila b sea más hidrófila (soluble en metanol al 92%) que la clorofila a (más soluble en éter). La gran mayoría de las plantas contienen dos o tres veces más clorofila a que clorofila b. La estructura de la molécula de clorofila fue determinada a principios de este siglo por los investigadores alemanes R. Wilstatter y A. Stoll y el famoso químico orgánico Hans Fischer.
Las clorofilas a y b absorben principalmente luz azul-violeta y roja, y
solo ligeramente luz verde. En el caso de la clorofila a, la máxima absorción se produce en el rango violeta, es decir, a longitudes de onda más cortas que en el caso de la clorofila b; mientras que en el rango rojo, el máximo se desplaza hacia longitudes de onda más largas. La posición del máximo de absorción depende del disolvente en el que se disuelven los colorantes, es decir, del tipo de moléculas de disolvente con las que se asocian las moléculas del colorante. Por ejemplo, en una planta viva, el pico de absorción principal de la clorofila a en el rango rojo se encuentra a 683 nm, que está desplazado en relación con el pico a 660 nm para esta clorofila disuelta en éter etílico. Para la clorofila a, también se encontró un pico de absorción adicional a 672 nm. Por lo tanto, se supone que esto es el resultado de condiciones ambientales ligeramente modificadas. Los gráficos que muestran la dependencia de la absorción de luz en diferentes longitudes de onda se denominan espectros de absorción. El espectro de absorción se determina utilizando un espectrofotómetro, es decir, un instrumento equipado con una fuente de luz que emite ondas de las longitudes de onda deseadas; un prisma o una red de difracción que sirve para dispersar la luz; espejos que reflejan longitudes de onda seleccionadas y las dirigen hacia la muestra que se está analizando, y un detector utilizado para detectar la cantidad de luz absorbida por la muestra que se está analizando. La solución de clorofila irradiada fluoresce emiten una intensa luz roja. Debido a que la mayor parte de la energía que causa la fluorescencia en las plantas vivas se utiliza en la fotosíntesis, no presentan fluorescencia visible.
3. Síntesis de clorofila.
La síntesis de clorofila depende de varios factores, como las características genéticas, la luz y la disponibilidad de ciertos minerales. En algunas variedades de maíz y girasol, algunas semillas germinadas producen plántulas sin clorofila. Este es un rasgo recesivo innato. Estos brotes, llamados albinos, mueren cuando se agotan las reservas nutricionales de la semilla. Sin embargo, si se les suministran azúcares como fuente de energía, estas plantas pueden seguir creciendo casi hasta la madurez.
La luz es esencial para la síntesis de clorofila en casi todas las angiospermas, aunque las coníferas, los musgos, algunos helechos y muchas algas también pueden producir este pigmento en la oscuridad. Las angiospermas que crecen en la oscuridad sintetizan pequeñas cantidades de un pigmento verde similar a la clorofila, pero su contenido es demasiado bajo para conferirles un color verde. Al exponerse a la luz, este pigmento desaparece, siendo reemplazado por clorofila a, cuya cantidad aumenta con el tiempo. Este pigmento que desaparece se denomina protoclorofila. Químicamente, se diferencia de la clorofila a únicamente por la ausencia de dos átomos de hidrógeno en la molécula. Además, absorbe la luz azul y roja de forma similar, presentando máximos de absorción en longitudes de onda de 450 nm y 650 nm. Estas longitudes de onda son las más efectivas en la conversión de protoclorofila a clorofila a. Sin embargo, el origen de la clorofila b sigue sin explicación.
Parece que varios elementos minerales participan directamente en la síntesis de clorofila, ya que su deficiencia provoca decoloración, o clorosis, en las plantas. Los anillos de pirrol de la clorofila se forman mediante la combinación previa de glicina con ácido succínico. Entre la reacción de estos dos compuestos y la formación final del compuesto complejo clorofila, se produce una serie de reacciones intermedias catalizadas por enzimas específicas. La reacción entre la glicina y el succinil-CoA depende de la presencia de hierro como coenzima, y una deficiencia de este elemento provoca clorosis rápida en las hojas jóvenes en desarrollo. Las deficiencias de manganeso, potasio, zinc y cobre también causan clorosis, pero aún no se comprende el papel de estos elementos. Presumiblemente, activan ciertas enzimas implicadas en la síntesis de clorofila. La deficiencia de manganeso conduce a la destrucción de la estructura lamelar interna de los cloroplastos y puede ser la razón de la inhibición de la síntesis de clorofila que estabiliza las lamelas.
Sin embargo, la falta de nitrógeno y magnesio provoca el amarillamiento de las plantas. La función de estos elementos es comprensible, ya que son los componentes estructurales básicos de la clorofila. Esto no excluye sus funciones indirectas; por ejemplo, es probable que el magnesio actúe como coenzima en una o más etapas de la síntesis de clorofila.
4. Estructura y síntesis de los pigmentos carotenoides.
Los carotenoides son un grupo de pigmentos que, en las plantas superiores, suelen ser rojos, naranjas, amarillos o marrones y se encuentran en los cloroplastos junto con la clorofila. También se encuentran a veces en otros plastidios, como los cromoplastos de las raíces de zanahoria y tomate, y en los cromoplastos de numerosas flores amarillas. El color amarillo de los carotenoides es visible en las angiospermas que crecen en la oscuridad y carecen de clorofila (lo que indica que la luz no es necesaria para su acumulación) y en otoño en las hojas de muchas especies, donde desaparece la clorofila. De hecho, los carotenoides también pueden presentar otros colores, como el verde en los aguacates, el rosa en los camarones o casi el negro tinta en los cefalópodos. Estos pigmentos se encuentran en todos los grupos principales de plantas y animales, pero nuestra discusión se centrará principalmente en aquellos que se encuentran en los cloroplastos junto con la clorofila. Los carotenoides se pueden dividir en dos grupos químicos: carotenos y xantofilas. Dentro de cada grupo, existen varios tipos distintos de estos compuestos, cuyas moléculas están compuestas por 40 átomos de carbono. Los carotenoides, cuyo representante más importante es el β-caroteno, son hidrocarburos (Fig. 2). Las xantofilas, como la luteína y la zeaxantina, tienen átomos de oxígeno unidos a sus anillos terminales. Tanto las xantofilas como los carotenos se sintetizan en las plantas a partir de acetato activo, es decir, acetil-CoA, a través del ácido mevalónico.
Figura 2. Fórmulas estructurales de los carotenoides.
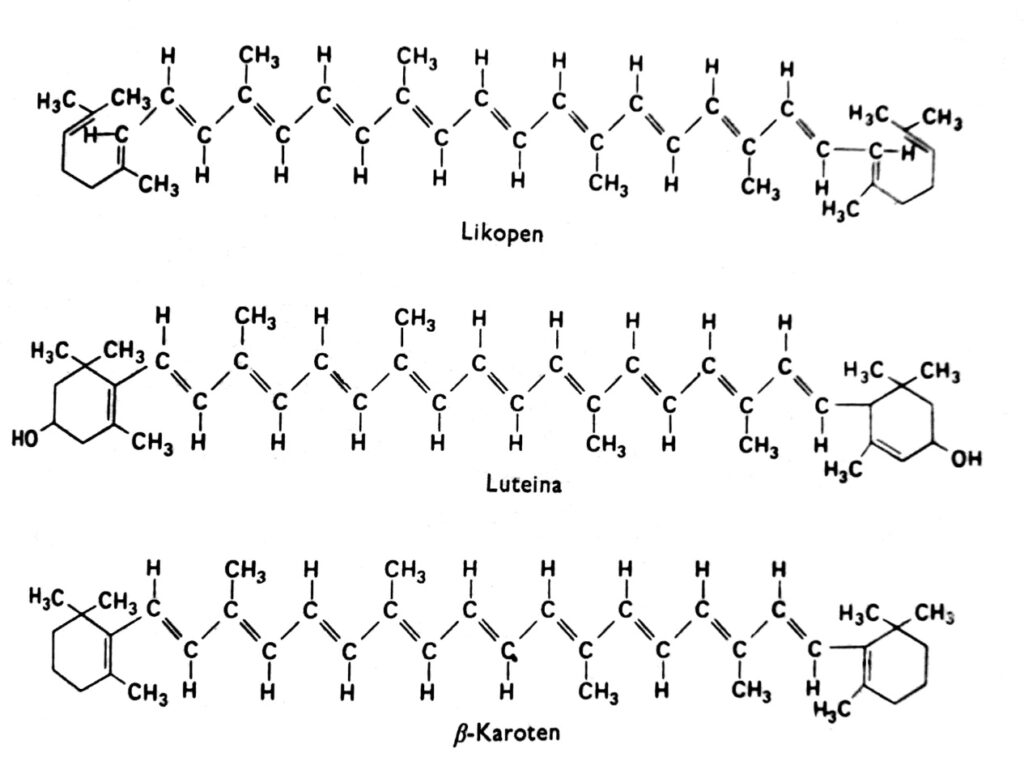
Casi ningún carotenoide se disuelve en agua, pero sí en muchos disolventes orgánicos, como la acetona y el éter. Las xantofilas, al tener átomos de oxígeno adicionales, son más hidrófilas que los carotenos. En solución, la mayoría de los carotenoides absorben principalmente luz violeta y azul en el rango de 400-500 nm. Sus espectros de absorción dependen del grado de saturación de los enlaces dobles y, por lo tanto, pueden ser ligeramente diferentes en cloroplastos intactos. Existe evidencia de que, en células intactas, los máximos de absorción de algunos carotenoides a 500 nm se desplazan a 450 nm después de la extracción de estos pigmentos con un disolvente orgánico.
5. Funciones de los carotenoides.
Los carotenoides desempeñan un papel importante en la fotosíntesis, probablemente por dos razones:
1. Cumplen una función protectora, protegiendo la clorofila de los efectos dañinos de la luz
y el oxígeno. Este fenómeno destructivo se denomina fotooxidación. A menudo se observa que una solución alcohólica de clorofila se decolora bajo la influencia de la luz, lo que se acompaña de un consumo de oxígeno. Presumiblemente, una reacción similar ocurriría normalmente en las plantas si los carotenoides no cumplieran su función protectora. La evidencia más convincente proviene de observaciones de ciertos mutantes bacterianos. Debido a que no contienen pigmentos carotenoides, la fotooxidación ocurre con mucha facilidad en ellos. Además, se conocen mutantes de girasol albinos que han perdido la capacidad de sintetizar carotenoides pero aún producen cantidades mínimas de clorofila. En este caso, el aumento de la intensidad de la luz desencadena una fotooxidación adicional de la clorofila, y la velocidad de este proceso es igual a su síntesis. El mecanismo por el cual los carotenoides protegen la clorofila de la oxidación aún no está claro. Sin embargo, se cree que los carotenoides se fotooxidan por la luz, lo que podría descomponer la clorofila.
2. Otra posible función de los carotenoides, y especialmente de las xantofilas, es su participación en la absorción de luz en longitudes de onda activas en la fotosíntesis. Existe evidencia irrefutable de que algunos carotenoides, como la fucoxantina (también llamada fucoxantol), presente en algas pardas (p. ej., fucus), participan activamente en la fotosíntesis. Esta evidencia se obtuvo comparando el espectro de absorción de la fucoxantina con el denominado espectro funcional, o espectro de actividad fotosintética. El espectro funcional fotosintético es un gráfico que ilustra la eficiencia de la fotosíntesis en función de las longitudes de onda de la luz que desencadenan este proceso. Se ha descubierto que en las plantas verdes y superiores, la luz absorbida por algunos carotenoides también se utiliza en la fotosíntesis. En estos casos, los máximos del espectro funcional se encuentran en los rangos de luz roja y azul, donde la clorofila absorbe con mayor intensidad. Sin embargo, la fotosíntesis también se produce utilizando luz azul-verde, presumiblemente porque algunos carotenoides son activos en este rango. La evidencia más convincente de que los carotenoides participan en la fotosíntesis es su capacidad para transferir energía de excitación directamente a la clorofila a. Cuando una planta se irradia con luz cuyas longitudes de onda son absorbidas casi exclusivamente por los carotenoides, se observa fluorescencia roja proveniente de la clorofila a. Se postula que la energía de excitación, a veces liberada como fluorescencia, se utiliza típicamente como fuerza impulsora de la fotosíntesis.
6. Ficobilinas.
El último grupo de pigmentos fotosintéticos son las ficobilinas, presentes en cianobacterias
y algas rojas. Estos pigmentos se clasifican en dos grupos: rojos (ficoeritrinas) y azules (ficocianinas). Al igual que la clorofila, están compuestos por cuatro anillos de pirrol. Sin embargo, las ficobilinas contienen un sistema de anillos de pirrol abierto y carecen de magnesio y de un grupo fitol. En las células vivas, se encuentran asociadas a proteínas y son solubles en agua. Ambos tipos de pigmentos absorben la luz, utilizándola para la fotosíntesis. En las algas rojas, la mayor eficiencia fotosintética se produce en el rango de luz verde, ya que en este rango, la ficoeritrina, a diferencia de la clorofila, absorbe la luz con mayor intensidad. Al igual que los carotenoides, las ficobilinas transfieren energía de excitación a la clorofila a y pueden provocar su fluorescencia. Este modo de transferencia de energía indica que las ficobilinas participan activamente en la fotosíntesis capturando fotones, cuya energía suele ser utilizada por la clorofila a como fuerza impulsora de la fotosíntesis. Los pigmentos como los carotenoides y las ficobilinas, que transfieren energía a un aceptor adecuado, por ejemplo, la clorofila a, se denominan pigmentos accesorios. El grupo de pigmentos accesorios también incluye las clorofilas b, c y d, ya que absorben energía lumínica, la cual, a su vez, puede transferirse a la clorofila a.
Literatura:
-Franck B. Salibury, Cleon Ross "Fisiología vegetal" PWRiL 1975.
-Otis F. Daniel G. Curtis Clark "Introducción a la fisiología vegetal" PWRiL 1958.
-Zurzycki J. Michniewicz M. "Fisiología vegetal" PWRiL 1979.
-Richter G. "Procesos metabólicos en las plantas" PWN 1975.