1. Einleitung.
Die Photosynthese findet in allen Algen und höheren Pflanzen in Gegenwart von Chlorophyll a; eine Ausnahme bilden photosynthetische Bakterien, die Pigmente mit einer leicht abweichenden Struktur als Chlorophyll a. Auch andere Chlorophyllvarianten kommen in Pflanzen vor.
Höhere Pflanzen und Grünalgen enthalten Chlorophyll a und b, während Cyanobakterien und einige Rotalgen nur Chlorophyll a. Andere Rotalgenarten enthalten Chlorophyll a und eine geringe Menge Chlorophyll d, und Braunalgen und Kieselalgen besitzen sowohl Chlorophyll aals auch Chlorophyll c.
Darüber hinaus sind weitere Pigmente an der Photosynthese beteiligt, wie beispielsweise Carotinoide und in Rotalgen und Cyanobakterien Phycobilin-Pigmente (Phycoerythrin und Phycocyanin). Tabelle 1 listet die in verschiedenen photosynthetischen Organismen vorkommenden Pigmente auf, einschließlich ihrer Farbe und ihres primären Absorptionsmaximums. Abbildung 1 zeigt die grundlegenden chemischen Strukturen einiger Photosynthese-Pigmente.
Abb. 1. Strukturformeln der Chlorophylle.
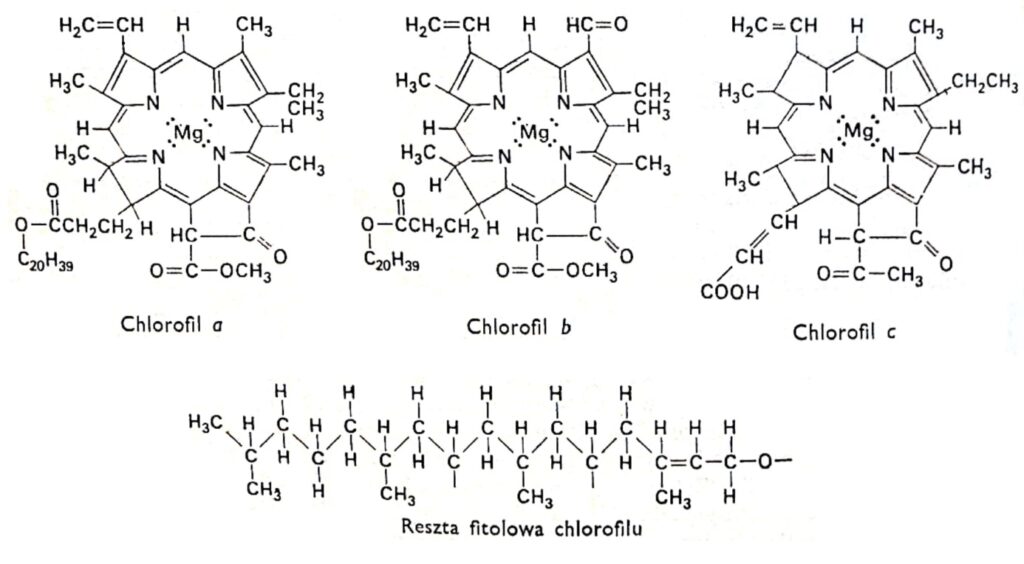
Tabelle 1. Eigenschaften der Photosynthesepigmente.
Farbstoff | Farbe | Hauptabsorptionsmaxima im sichtbaren Bereich (nm) |
| Chlorophyll a | blaugrün | 430 (blauviolett) 660 (rot) |
| Chlorophyll b | Grün | 455 (blau) 640 (orange-rot) |
| Phycocyanine | Blau | 560-660 (gelb bis orangerot) |
| Phycoerythrine | Rot | 550-570 (grün bis gelb) |
| Carotinoide | Gelb, orangerot | 430-490 (violett bis blaugrün) |
2. Die Struktur des Chlorophylls.
Alle Chlorophyllarten besitzen die gleiche Porphyrinstruktur, bestehend aus vier Pyrrolringen, die über Stickstoffatome an Magnesium gebunden sind. Das Chlorophyllmolekül enthält außerdem einen fünften Ring, der vollständig aus Kohlenstoffatomen besteht und mit dem langkettigen Alkohol Phytol verbunden ist, der typisch für die meisten Chlorophylle ist, mit Ausnahme von Chlorophyll c. Chlorophylle unterscheiden sich in der Struktur der an die Pyrrolringe gebundenen Seitenketten. Der Unterschied zwischen Chlorophyll a und b besteht darin, dass der dritte Ring von Chlorophyll a eine Methylgruppe, der von Chlorophyll b eine Aldehydgruppe enthält. Die Aldehydgruppe macht Chlorophyll b hydrophiler (löslich in 92%igem Methanol) als Chlorophyll a (besser löslich in Ether). Die überwiegende Mehrheit der Pflanzen enthält zwei- bis dreimal so viel Chlorophyll a wie Chlorophyll b. Die Struktur des Chlorophyllmoleküls wurde zu Beginn dieses Jahrhunderts von den deutschen Forschern R. Wilstatter und A. Stoll sowie dem berühmten organischen Chemiker Hans Fischer aufgeklärt.
Chlorophyll a und b absorbieren hauptsächlich blauviolettes und rotes Licht und
nur geringfügig grünes Licht. Bei Chlorophyll a liegt das Absorptionsmaximum im violetten Bereich, also bei kürzeren Wellenlängen als bei Chlorophyll b; im roten Bereich ist es hingegen zu längeren Wellenlängen verschoben. Die Lage des Absorptionsmaximums hängt vom Lösungsmittel ab, in dem die Farbstoffe gelöst sind, d. h. von der Art der Lösungsmittelmoleküle, mit denen die Farbstoffmoleküle assoziiert sind. Beispielsweise liegt in einer lebenden Pflanze das Hauptabsorptionsmaximum von Chlorophyll a im roten Bereich bei 683 nm, was im Vergleich zum Maximum bei 660 nm für in Diethylether gelöstes Chlorophyll verschoben ist. Für Chlorophyll a wurde außerdem ein zusätzliches Absorptionsmaximum bei 672 nm gefunden. Es wird daher angenommen, dass dies auf leicht veränderte Umgebungsbedingungen zurückzuführen ist. Diagramme, die die Abhängigkeit der Lichtabsorption von verschiedenen Wellenlängen darstellen, werden als Absorptionsspektren bezeichnet. Das Absorptionsspektrum wird mithilfe eines Spektralphotometers bestimmt, d. h. eines Geräts, das mit einer Lichtquelle ausgestattet ist, die Wellen der gewünschten Wellenlängen aussendet; einem Prisma oder einem Beugungsgitter zur Lichtstreuung; Spiegeln, die ausgewählte Wellenlängen reflektieren und auf die zu untersuchende Probe lenken; und einem Detektor zur Messung der von der Probe absorbierten Lichtmenge. Die bestrahlte Chlorophylllösung Sie fluoreszieren stark und emittieren intensives rotes Licht. Da der größte Teil der Energie, die bei lebenden Pflanzen Fluoreszenz verursacht, für die Photosynthese verwendet wird, fluoreszieren sie nicht sichtbar.
3. Chlorophyllsynthese.
Die Chlorophyllsynthese hängt von verschiedenen Faktoren ab, darunter genetische Merkmale, Licht und die Verfügbarkeit bestimmter Mineralstoffe. Bei manchen Mais- und Sonnenblumensorten keimen einige Samen und bringen Sämlinge hervor, denen Chlorophyll fehlt. Dies ist ein angeborenes rezessives Merkmal. Diese Triebe, sogenannte Albinos, sterben ab, sobald die Nährstoffreserven des Samens aufgebraucht sind. Werden diese Pflanzen jedoch mit Zucker als Energiequelle versorgt, können sie bis fast zur Reife weiterwachsen.
Licht ist für die Chlorophyllsynthese in fast allen Angiospermen unerlässlich, obwohl Nadelbäume, Moose, einige Farne und viele Algen dieses Pigment auch im Dunkeln produzieren können. Im Dunkeln wachsende Angiospermen synthetisieren geringe Mengen eines grünen, dem Chlorophyll ähnlichen Pigments, dessen Konzentration jedoch zu gering ist, um den Pflanzen eine grüne Farbe zu verleihen. Bei Lichteinwirkung verschwindet dieses Pigment und wird durch Chlorophyll a ersetzt , dessen Menge mit der Zeit zunimmt. Dieses verschwindende Pigment wird Protochlorophyll genannt. Chemisch unterscheidet es sich von Chlorophyll a lediglich durch das Fehlen zweier Wasserstoffatome im Molekül. Darüber hinaus absorbiert es blaues und rotes Licht in ähnlicher Weise und weist Absorptionsmaxima bei Wellenlängen von 450 nm und 650 nm auf. Diese Wellenlängen sind am effektivsten für die Umwandlung von Protochlorophyll in Chlorophyll a . Der Ursprung von Chlorophyll b ist jedoch weiterhin ungeklärt.
Es scheint, dass mehrere Mineralstoffe direkt an der Chlorophyllsynthese beteiligt sind, da ihr Mangel zu Verfärbungen (Chlorose) in Pflanzen führt. Die Pyrrolringe des Chlorophylls entstehen durch die vorherige Verbindung von Glycin mit Bernsteinsäure. Zwischen der Reaktion dieser beiden Verbindungen und der endgültigen Bildung des komplexen Chlorophylls finden eine Reihe von Zwischenreaktionen statt, die von spezifischen Enzymen katalysiert werden. Die Reaktion zwischen Glycin und Succinyl-CoA ist abhängig von Eisen als Coenzym, und ein Mangel an diesem Element verursacht eine rasche Chlorose in jungen, sich entwickelnden Blättern. Auch ein Mangel an Mangan, Kalium, Zink und Kupfer führt zu Chlorose, die Rolle dieser Elemente ist jedoch noch nicht vollständig geklärt. Vermutlich aktivieren sie bestimmte Enzyme, die an der Chlorophyllsynthese beteiligt sind. Manganmangel führt zur Zerstörung der inneren Lamellenstruktur der Chloroplasten und könnte die Ursache für die Hemmung der Chlorophyllsynthese sein, wodurch die Lamellen stabilisierende Chlorophyllbildung gehemmt wird.
Ein Mangel an Stickstoff und Magnesium führt jedoch zur Vergilbung der Pflanzen. Die Rolle dieser Elemente ist nachvollziehbar, da sie die grundlegenden Bausteine des Chlorophylls sind. Dies schließt jedoch indirektere Funktionen nicht aus; so wirkt Magnesium beispielsweise wahrscheinlich als Coenzym in einem oder mehreren Schritten der Chlorophyllsynthese.
4. Struktur und Synthese der Carotinoidpigmente.
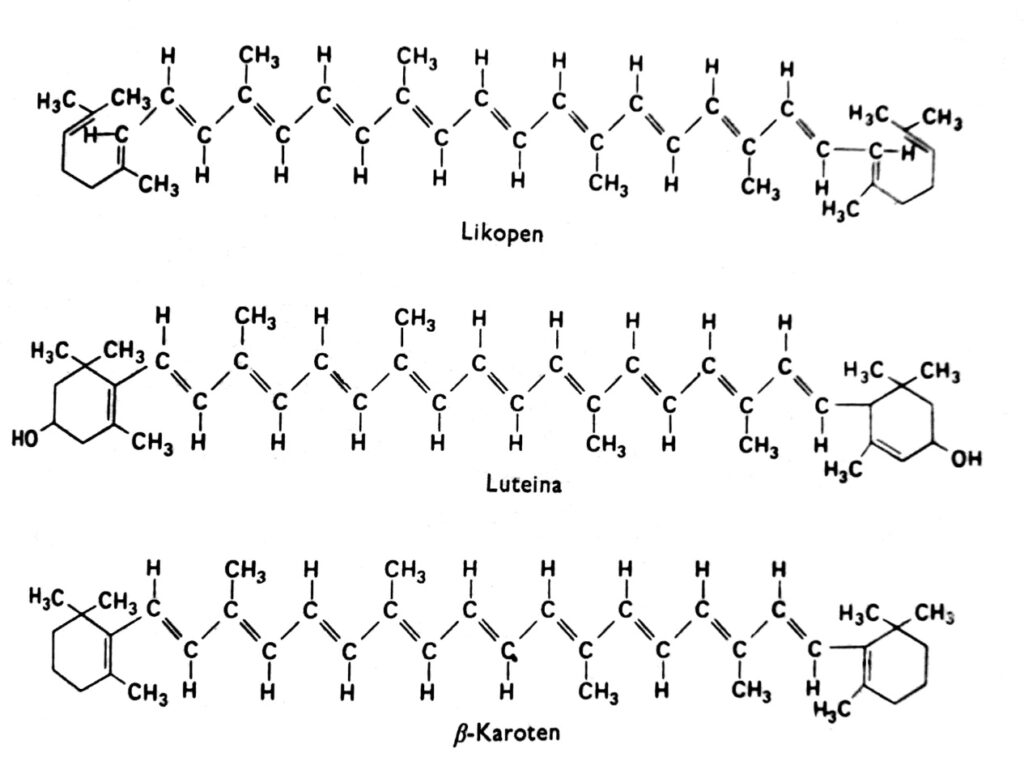
Carotinoide sind eine Gruppe von Pigmenten, die in höheren Pflanzen im Allgemeinen rot, orange, gelb oder braun sind und in Chloroplasten zusammen mit Chlorophyll vorkommen. Sie finden sich mitunter auch in anderen Plastiden, beispielsweise in den Chromoplasten von Karotten- und Tomatenwurzeln sowie in den Chromoplasten zahlreicher gelber Blüten. Die gelbe Farbe der Carotinoide ist bei Angiospermen sichtbar, die im Dunkeln wachsen und kein Chlorophyll besitzen (was darauf hindeutet, dass Licht für ihre Anreicherung nicht notwendig ist), und im Herbst in den Blättern vieler Arten, in denen das Chlorophyll abgebaut wird. Tatsächlich können Carotinoide auch andere Farben annehmen, wie Grün in Avocados, Rosa in Garnelen oder fast Tintenschwarz in Kopffüßern. Diese Pigmente kommen in allen wichtigen Pflanzen- und Tiergruppen vor, unsere Betrachtung konzentriert sich jedoch hauptsächlich auf die in Chloroplasten zusammen mit Chlorophyll vorkommenden Carotinoide. Man unterscheidet zwei chemische Gruppen von Carotinoiden: Carotine und Xanthophylle. Innerhalb jeder Gruppe existieren mehrere verschiedene Arten dieser Verbindungen, deren Moleküle aus 40 Kohlenstoffatomen bestehen. Carotinoide, deren wichtigster Vertreter β-Carotin ist, sind Kohlenwasserstoffe (Abb. 2). Xanthophylle wie Lutein und Zeaxanthin besitzen Sauerstoffatome an ihren endständigen Ringen. Sowohl Xanthophylle als auch Carotine werden in Pflanzen aus aktivem Acetat, d. h. Acetyl-CoA, über Mevalonsäure synthetisiert.
Abb. 2. Strukturformeln der Carotinoide.
Fast keines der Carotinoide ist in Wasser löslich, wohl aber in vielen organischen Lösungsmitteln wie Aceton und Ether. Xanthophylle, die zusätzliche Sauerstoffatome besitzen, sind hydrophiler als Carotine. In Lösung absorbieren die meisten Carotinoide vorwiegend violettes und blaues Licht im Bereich von 400–500 nm. Ihre Absorptionsspektren hängen vom Sättigungsgrad der Doppelbindungen ab und können daher in intakten Chloroplasten leicht abweichen. Es gibt Hinweise darauf, dass sich in intakten Zellen die Absorptionsmaxima einiger Carotinoide von 500 nm nach Extraktion dieser Pigmente mit einem organischen Lösungsmittel zu 450 nm verschieben.
5. Funktionen der Carotinoide.
Carotinoide spielen eine wichtige Rolle bei der Photosynthese, wahrscheinlich aus zwei Gründen:
1. Carotinoide üben eine Schutzfunktion aus, indem sie Chlorophyll vor den schädlichen Auswirkungen von Licht
und Sauerstoff bewahren. Dieses zerstörerische Phänomen wird als Photooxidation bezeichnet. Häufig beobachtet man, dass sich eine alkoholische Chlorophylllösung unter Lichteinwirkung entfärbt, was mit Sauerstoffverbrauch einhergeht. Vermutlich würde eine ähnliche Reaktion normalerweise auch in Pflanzen auftreten, wenn Carotinoide ihre Schutzfunktion nicht erfüllen würden. Die überzeugendsten Beweise stammen von Beobachtungen an bestimmten Bakterienmutanten. Da diese keine Carotinoidpigmente enthalten, findet in ihnen die Photooxidation sehr schnell statt. Darüber hinaus sind Albino-Sonnenblumenmutanten bekannt, die die Fähigkeit zur Carotinoidsynthese verloren haben, aber dennoch Spuren von Chlorophyll produzieren. In diesem Fall löst eine erhöhte Lichtintensität eine zusätzliche Photooxidation des Chlorophylls aus, deren Geschwindigkeit der Syntheserate entspricht. Der Mechanismus, durch den Carotinoide Chlorophyll vor Oxidation schützen, ist noch nicht vollständig geklärt. Es wird jedoch angenommen, dass Carotinoide durch Licht photooxidiert werden, wodurch Chlorophyll abgebaut werden könnte.
2. Eine weitere wahrscheinliche Funktion von Carotinoiden, insbesondere von Xanthophyllen, ist ihre Beteiligung an der Absorption von Licht in Wellenlängenbereichen, die für die Photosynthese relevant sind. Es gibt unbestreitbare Belege dafür, dass einige Carotinoide, wie beispielsweise Fucoxanthin (auch Fucoxanthol genannt), das in Braunalgen (z. B. Fucus) vorkommt, aktiv an der Photosynthese beteiligt sind. Diese Belege wurden durch den Vergleich des Absorptionsspektrums von Fucoxanthin mit dem sogenannten funktionellen Spektrum, also dem Spektrum der photosynthetischen Aktivität, gewonnen. Das photosynthetische funktionelle Spektrum ist ein Diagramm, das die Effizienz der Photosynthese in Abhängigkeit von den Wellenlängen des Lichts darstellt, die diesen Prozess auslösen. Es wurde festgestellt, dass in grünen Pflanzen und höheren Pflanzen das von einigen Carotinoiden absorbierte Licht ebenfalls für die Photosynthese genutzt wird. In solchen Fällen liegen die Maxima des funktionellen Spektrums im roten und blauen Lichtbereich, wo Chlorophyll am stärksten absorbiert. Dennoch findet Photosynthese auch mit blaugrünem Licht statt, vermutlich weil einige Carotinoide in diesem Bereich aktiv sind. Der überzeugendste Beweis für die Beteiligung von Carotinoiden an der Photosynthese ist ihre Fähigkeit, Anregungsenergie direkt auf Chlorophyll a zu übertragen. Wird eine Pflanze mit Licht bestrahlt, dessen Wellenlängen fast ausschließlich von Carotinoiden absorbiert werden, ist eine rote Fluoreszenz des Chlorophyll a zu beobachten. Es wird angenommen, dass die Anregungsenergie, die mitunter als Fluoreszenz freigesetzt wird, typischerweise als treibende Kraft für die Photosynthese dient.
6. Phycobiline.
Die letzte Gruppe der Photosynthesepigmente bilden die Phycobiline, die in Cyanobakterien
und Rotalgen vorkommen. Diese Pigmente werden in zwei Gruppen unterteilt: rote (Phycoerythrine) und blaue (Phycocyanine). Wie Chlorophyll bestehen sie aus vier Pyrrolringen. Phycobiline besitzen jedoch ein offenes Pyrrolringsystem und weder Magnesium noch eine Phytolgruppe. In lebenden Zellen kommen sie in Verbindung mit Proteinen vor und sind wasserlöslich. Beide Pigmenttypen absorbieren Licht und nutzen es für die Photosynthese. Bei Rotalgen ist die Photosynthese im grünen Lichtbereich am effizientesten, da Phycoerythrin in diesem Bereich – anders als Chlorophyll – Licht am intensivsten absorbiert. Wie Carotinoide übertragen Phycobiline Anregungsenergie auf Chlorophyll a und können dieses zur Fluoreszenz anregen. Diese Art des Energietransfers deutet darauf hin, dass Phycobiline aktiv an der Photosynthese beteiligt sind, indem sie Photonen einfangen. Die Energie dieser Photonen wird typischerweise von Chlorophyll a als Antriebskraft für die Photosynthese genutzt. Pigmente wie Carotinoide und Phycobiline, die Energie an einen geeigneten Akzeptor, z. B. Chlorophyll a, abgeben, werden als akzessorische Pigmente bezeichnet. Zu den akzessorischen Pigmenten gehören auch die Chlorophylle b, c und d, da diese Lichtenergie absorbieren, die wiederum an Chlorophyll a.
Literatur:
-Franck B. Salibury, Cleon Ross "Plant Physiology" PWRiL 1975.
-Otis F. Daniel G. Curtis Clark "Einführung in die Pflanzenphysiologie" PWRiL 1958.
-Zurzycki J. Michniewicz M. „Plant Physiology“ PWRiL 1979.
-Richter G. „Metabolische Prozesse in Pflanzen“ PWN 1975.