1.引言。.
所有藻类和高等植物的光合作用都发生在叶绿素 a;例外的是光合细菌,它们含有与叶绿素 a。植物中也存在其他类型的叶绿素。
高等植物和绿藻含有叶绿素 a 和 b,而蓝藻和一些红藻只含有叶绿素 a。其他红藻种类含有叶绿素 a 和少量叶绿素 d,褐藻和硅藻则同时含有叶绿素 a和叶绿素 c。
此外,其他色素也参与光合作用,例如类胡萝卜素;在红藻和蓝藻中,还存在藻胆色素(藻红蛋白和藻蓝蛋白)。表1列出了各种光合生物中发现的色素,包括它们的颜色和主要吸收峰。图1展示了一些光合色素的基本化学结构。.
图1.叶绿素的结构式。.
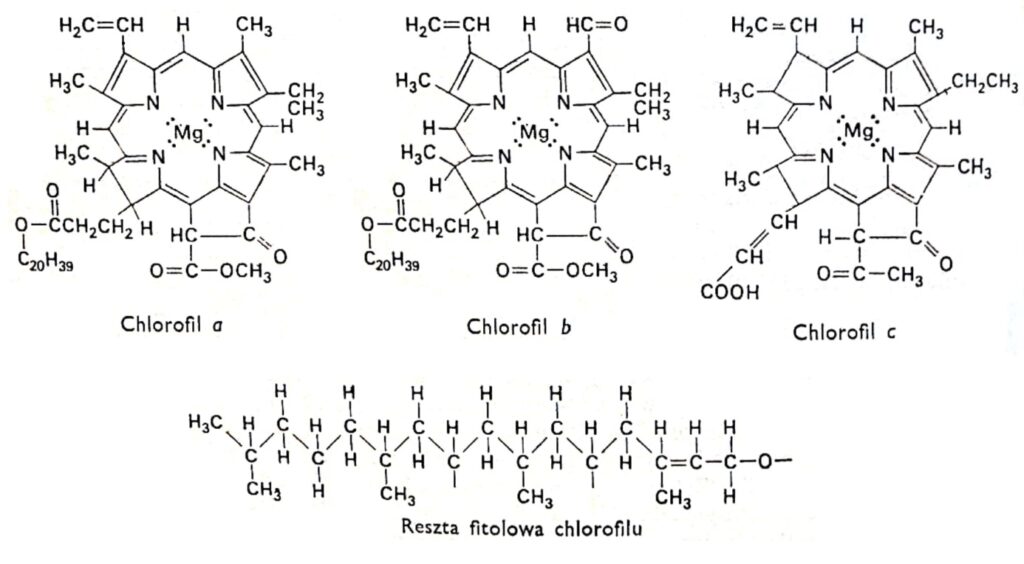
表1. 光合色素的特性。.
染料 | 颜色 | 可见光范围内的主要吸收峰 (nm) |
| 叶绿素 a | 蓝绿色 | 430(蓝紫色)660(红色) |
| 叶绿素 b | 绿色的 | 455(蓝色)640(橙红色) |
| 藻蓝蛋白 | 蓝色的 | 560-660(黄色至橙红色) |
| 藻红蛋白 | 红色的 | 550-570(绿转黄) |
| 类胡萝卜素 | 黄色、橙红色 | 430-490(紫色至蓝绿色) |
2. 叶绿素的结构。.
所有叶绿素都具有相同的卟啉结构,由四个吡咯环通过氮原子与镁原子连接而成。此外,叶绿素分子还包含一个完全由碳原子构成的第五环,该环与长链醇植醇相连,这是大多数叶绿素的特征,叶绿素c除外。叶绿素之间的区别在于与吡咯环相连的侧链结构不同。叶绿素a和b的区别在于,叶绿素a的第三个环含有一个甲基,而叶绿素b的第三个环含有一个醛基。醛基的存在使得叶绿素b比叶绿素a更具亲水性(更易溶于92%甲醇,而叶绿素a更易溶于乙醚)。绝大多数植物中叶绿素a的含量是叶绿素b的两到三倍。本世纪初,德国研究人员 R. Wilstatter 和 A. Stoll 以及著名有机化学家汉斯·费舍尔确定了叶绿素分子的结构。
叶绿素 a 和 b 主要吸收蓝紫光和红光,
仅少量吸收绿光。叶绿素a的最大吸收波长位于紫光波段,即比叶绿素 b;而叶绿素a在红光波段的最大吸收波长则更长。吸收峰的位置取决于染料溶解的溶剂,即染料分子所结合的溶剂分子类型。例如,在活体植物中,叶绿素 a 在红色波段的主要吸收峰位于683 nm处,相对于溶解在乙醚中的叶绿素a的吸收峰(660 nm)发生了偏移。此外 ,还发现了叶绿素a 在672 nm处的另一个吸收峰。因此,推测这是环境条件略微变化的结果。显示光吸收与不同波长关系的图表称为吸收光谱。吸收光谱的测定使用分光光度计,即一种配备有光源(发射所需波长的光)、棱镜或衍射光栅(用于色散光)、反射特定波长并将其导向待测样品的反射镜以及检测器(用于检测待测样品吸收的光量)的仪器。叶绿素溶液会发出强烈的荧光,发出耀眼的红光。由于活体植物产生荧光所需的大部分能量都用于光合作用,因此它们本身不会发出肉眼可见的荧光。
3. 叶绿素合成。.
叶绿素的合成取决于多种因素,包括遗传特性、光照以及某些矿质元素的供应。在某些玉米和向日葵品种中,部分发芽的种子会发育成缺乏叶绿素的幼苗。这是一种先天性隐性性状。这些被称为白化植株的幼苗会在种子营养储备耗尽时死亡。然而,如果为这些植株提供糖类作为能量来源,它们可以继续生长直至接近成熟。.
光照对于几乎所有被子植物合成叶绿素都至关重要,尽管针叶树、苔藓、一些蕨类植物和许多藻类也能在黑暗中合成这种色素。在黑暗中生长的被子植物会合成少量类似于叶绿素的绿色色素,但其含量太低,不足以使这些植物呈现绿色。一旦暴露在光照下,这种色素就会消失,取而代之的是叶绿素a ,其含量会随时间增加。这种消失的色素被称为原叶绿素。从化学结构上看,它与叶绿素a的区别仅在于分子中缺少两个氢原子。此外,它对蓝光和红光的吸收相似,在450纳米和650纳米波长处具有吸收峰值。这些波长的光最能有效地将原叶绿素转化为叶绿素a 。然而,叶绿素b的起源至今仍未阐明。
多种矿质元素似乎直接参与叶绿素的合成,因为它们的缺乏会导致植物褪色,即黄化病。叶绿素吡咯环是由甘氨酸与琥珀酸预先结合形成的。在这两种化合物反应到最终形成复杂的叶绿素化合物之间,会发生一系列由特定酶催化的中间反应。甘氨酸与琥珀酰辅酶A的反应依赖于铁作为辅酶的存在,铁的缺乏会导致幼嫩发育中的叶片迅速黄化。锰、钾、锌和铜的缺乏也会导致黄化病,但这些元素的作用机制尚不清楚。它们可能激活参与叶绿素合成的某些酶。锰缺乏会导致叶绿体内部层状结构的破坏,这可能是抑制叶绿素合成、从而稳定叶绿体层状结构的原因。.
然而,氮和镁的缺乏会导致植物叶片发黄。这些元素的作用不难理解,因为它们是叶绿素的基本结构成分。但这并不排除它们可能具有更间接的功能,例如,镁很可能在叶绿素合成的一个或多个步骤中作为辅酶发挥作用。.
4. 类胡萝卜素色素的结构和合成 。
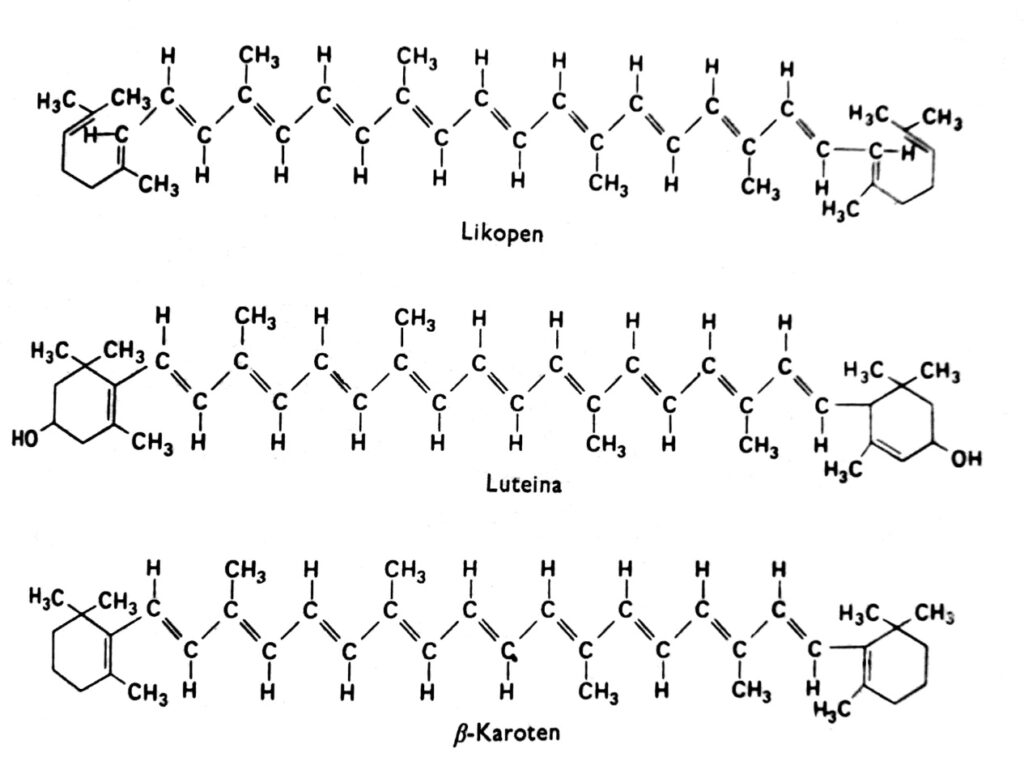
类胡萝卜素是一类色素,在高等植物中通常呈红色、橙色、黄色或棕色,与叶绿素一起存在于叶绿体中。它们有时也存在于其他质体中,例如胡萝卜和番茄根部的色素体,以及许多黄色花朵的色素体中。在黑暗中生长且缺乏叶绿素的被子植物中(这表明光照并非其积累的必要条件),以及在许多植物的秋季叶片中(此时叶绿素消失),类胡萝卜素的黄色清晰可见。事实上,类胡萝卜素还可以呈现其他颜色,例如鳄梨中的绿色、虾中的粉红色,或头足类动物中的近乎墨黑色。这些色素存在于所有主要的动植物类群中,但我们的讨论将主要集中在与叶绿素一起存在于叶绿体中的类胡萝卜素上。类胡萝卜素可分为两大类:胡萝卜素和叶黄素。每类化合物中都包含几种不同类型的化合物,它们的分子由40个碳原子组成。类胡萝卜素,其中最重要的代表是β-胡萝卜素,是碳氢化合物(图2)。叶黄素,例如叶黄素和玉米黄素,其末端环上连接着氧原子。叶黄素和胡萝卜素均由植物中的活性乙酸(即乙酰辅酶A)经甲羟戊酸合成。.
图 2. 类胡萝卜素的结构式。.
几乎所有类胡萝卜素都不溶于水,但它们能溶于许多有机溶剂,例如丙酮和乙醚。叶黄素由于含有额外的氧原子,比胡萝卜素更具亲水性。在溶液中,大多数类胡萝卜素主要吸收400-500纳米范围内的紫光和蓝光。它们的吸收光谱取决于双键的饱和度,因此在完整的叶绿体中可能略有不同。有证据表明,在完整细胞中,某些类胡萝卜素在500纳米处的吸收峰值在用有机溶剂提取后会移动到450纳米处。.
5. 类胡萝卜素的功能。.
类胡萝卜素在光合作用中发挥着重要作用,这可能有两个原因:
1. 类胡萝卜素发挥保护作用,保护叶绿素免受光和氧的损害
。这种破坏性现象称为光氧化。人们经常观察到,叶绿素的醇溶液在光照下会褪色,并伴随氧气的消耗。可以推测,如果类胡萝卜素不发挥其保护作用,植物中也会发生类似的反应。最有说服力的证据来自对某些细菌突变体的观察。由于这些细菌不含类胡萝卜素色素,因此它们很容易发生光氧化。此外,已知一些白化向日葵突变体丧失了合成类胡萝卜素的能力,但仍能产生微量的叶绿素。在这种情况下,光照强度的增加会引发叶绿素的进一步光氧化,并且该过程的速率与其合成速率相当。类胡萝卜素保护叶绿素免受氧化的机制尚不清楚。然而,人们认为类胡萝卜素会被光氧化,而光氧化可能会分解叶绿素。
2. 类胡萝卜素,尤其是叶黄素,可能具有的另一个功能是参与吸收光合作用活跃波长的光。有确凿的证据表明,某些类胡萝卜素,例如褐藻(如墨角藻)中的岩藻黄素(也称岩藻黄醇),能够积极参与光合作用。这一证据是通过比较岩藻黄素的吸收光谱与所谓的功能光谱(或称光合作用活性光谱)获得的。光合作用功能光谱是一个图表,它展示了光合作用效率与触发该过程的光波长之间的关系。研究发现,在绿色植物和高等植物中,某些类胡萝卜素吸收的光也被用于光合作用。在这种情况下,功能光谱的峰值落在红光和蓝光范围内,而叶绿素在这些光波段的吸收最强。然而,光合作用仍然利用蓝绿光进行,这可能是因为某些类胡萝卜素在该波段具有活性。类胡萝卜素参与光合作用的最有力证据是它们能够将激发能直接传递给叶绿素a。当植物被几乎完全被类胡萝卜素吸收的光波长照射时,会观察到叶绿素a发出的红色荧光。据推测,这种激发能(有时以荧光的形式释放)通常被用作光合作用的驱动力。.
6. 藻胆蛋白。.
最后一类光合色素是藻胆蛋白,存在于蓝藻
和红藻中。这些色素分为两类:红色(藻红蛋白)和蓝色(藻蓝蛋白)。与叶绿素一样,它们也由四个吡咯环组成。然而,藻胆蛋白的吡咯环是开放的,并且不含镁或植醇部分。在活细胞中,它们与蛋白质结合,并且溶于水。这两种色素都能吸收光能,用于光合作用。在红藻中,光合作用效率最高的是绿光波段,因为在这个波段,藻红蛋白(与叶绿素不同)对光的吸收最强。与类胡萝卜素一样,藻胆蛋白可以将激发态能量传递给叶绿素a,使其发出荧光。这种能量传递方式表明,藻胆蛋白通过捕获光子积极参与光合作用,其能量通常被叶绿素 a 用作光合作用的驱动力。类胡萝卜素和藻胆蛋白等色素能够将能量传递给合适的受体,例如叶绿素 a,这类色素被称为辅助色素。辅助色素还包括叶绿素 b、 c 和 d,因为它们能够吸收光能,并将能量传递给叶绿素 a。
文学:
-Franck B. Salibury,Cleon Ross《植物生理学》,PWRiL 1975。.
-Otis F. Daniel G. Curtis Clark《植物生理学导论》PWRiL 1958。.
-Zurzycki J. Michniewicz M.“植物生理学”PWRiL 1979。.
-Richter G.“植物代谢过程”PWN 1975。.