1. Введение.
Фотосинтез у всех водорослей и высших растений происходит в присутствии хлорофилла а; исключением являются фотосинтезирующие бактерии, которые содержат пигменты с несколько иной структурой, чем хлорофилл а. Другие разновидности хлорофилла также встречаются в растениях.
Высшие растения и зелёные водоросли содержат хлорофилл а и b, тогда как цианобактерии и некоторые красные водоросли содержат только хлорофилл а. Другие виды красных водорослей содержат хлорофилл а и небольшое количество хлорофилла d, а бурые водоросли и диатомовые водоросли содержат как хлорофилл а,так и хлорофилл с.
Кроме того, в фотосинтезе участвуют и другие пигменты, такие как каротиноиды, а в красных водорослях и цианобактериях — фикобилиновые пигменты (фикоэритрин и фикоцианин). В таблице 1 перечислены пигменты, обнаруженные в различных фотосинтезирующих организмах, включая их цвет и основной максимум поглощения. На рисунке 1 показаны основные химические структуры некоторых фотосинтетических пигментов.
Рис. 1. Структурные формулы хлорофиллов.
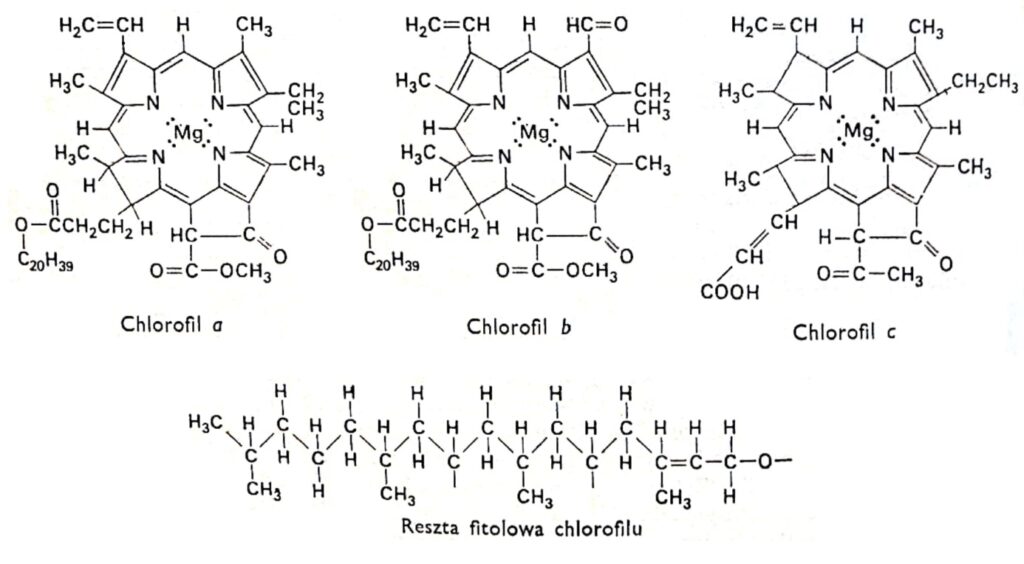
Таблица 1. Свойства фотосинтетических пигментов.
Краситель | Цвет | Основные пики поглощения в видимом диапазоне (нм) |
| Хлорофилл а | сине-зеленый | 430 (сине-фиолетовый) 660 (красный) |
| Хлорофилл b | зеленый | 455 (синий) 640 (оранжево-красный) |
| Фикоцианины | синий | 560-660 (от жёлтого до оранжево-красного) |
| Фикоэритрины | красный | 550-570 (от зеленого до желтого) |
| Каротиноиды | Жёлтый, оранжево-красный | 430-490 (от фиолетового до сине-зеленого) |
2. Структура хлорофилла.
Все разновидности хлорофилла имеют одинаковую порфириновую структуру, состоящую из четырех пиррольных колец, связанных с магнием через атомы азота. Кроме того, молекула хлорофилла содержит пятое кольцо, полностью состоящее из атомов углерода и связанное с длинноцепочечным спиртом фитолом, что характерно для большинства хлорофиллов, за исключением хлорофилла c. Хлорофиллы различаются структурой боковых цепей, присоединенных к пиррольным кольцам. Разница между хлорофиллом a и b заключается в том, что третье кольцо хлорофилла a содержит метильную группу, тогда как в хлорофилле b оно содержит альдегидную группу. Альдегидная группа делает хлорофилл b более гидрофильным (растворимым в 92% метиловом спирте), чем хлорофилл a (более растворимым в эфире). Подавляющее большинство растений содержат в два-три раза больше хлорофилла a, чем хлорофилла b. Структура молекулы хлорофилла была определена в начале этого столетия немецкими исследователями Р. Вильштеттером и А. Штоллом, а также известным химиком-органиком Хансом Фишером.
Хлорофиллы а и b поглощают преимущественно сине-фиолетовый и красный свет, и
лишь незначительно зеленый. Для хлорофилла а максимальное поглощение наблюдается в фиолетовом диапазоне, то есть на более коротких длинах волн, чем для хлорофилла b; тогда как в красном диапазоне максимум смещен в сторону более длинных волн. «Положение максимума поглощения зависит от растворителя, в котором растворены красители, то есть от типа молекул растворителя, с которыми связаны молекулы красителя. Например, в живом растении основной пик поглощения хлорофилла а в красном диапазоне находится на 683 нм, который смещен относительно пика на 660 нм для этого хлорофилла, растворенного в этиловом эфире. Для хлорофилла а также был обнаружен дополнительный пик поглощения на 672 нм. Поэтому предполагается, что это результат незначительного изменения условий окружающей среды. Графики, показывающие зависимость поглощения света от различных длин волн, называются спектрами поглощения. Спектр поглощения определяется с помощью спектрофотометра, то есть прибора, оснащенного источником света, излучающим волны желаемых длин волн; призмой или дифракционной решеткой, служащей для рассеивания света; зеркалами, отражающими выбранные длины волн и направляющими их на исследуемый образец, и детектором, используемым для определения количества света, поглощенного исследуемым образцом. Облученный раствор хлорофилла Растения сильно флуоресцируют, излучая интенсивный красный свет. Поскольку большая часть энергии, вызывающей флуоресценцию в живых растениях, расходуется на фотосинтез, они не флуоресцируют видимым образом.
3. Синтез хлорофилла.
Синтез хлорофилла зависит от ряда факторов, включая генетические характеристики, свет и наличие определенных минеральных элементов. У некоторых сортов кукурузы и подсолнечника некоторые прорастающие семена дают всходы, лишенные хлорофилла. Это врожденный рецессивный признак. Эти побеги, называемые альбиносами, погибают, когда истощаются питательные запасы семени. Однако, если эти растения получают сахара в качестве источника энергии, они могут продолжать расти почти до зрелости.
Свет необходим для синтеза хлорофилла почти у всех покрытосеменных растений, хотя хвойные, мхи, некоторые папоротники и многие водоросли также могут производить этот пигмент в темноте. Покрытосеменные, растущие в темноте, синтезируют небольшое количество зеленого пигмента, похожего на хлорофилл, но его содержание слишком мало, чтобы придать этим растениям зеленый цвет. При воздействии света этот пигмент исчезает, заменяясь хлорофиллом а, количество которого со временем увеличивается. Этот исчезающий пигмент называется протохлорофиллом. Химически он отличается от хлорофилла а только отсутствием двух атомов водорода в молекуле. Кроме того, он одинаково поглощает синий и красный свет, демонстрируя максимумы поглощения на длинах волн 450 нм и 650 нм. Эти длины волн света наиболее эффективны для превращения протохлорофилла в хлорофилл а. Однако происхождение хлорофилла b остается невыясненным.
По-видимому, ряд минеральных элементов непосредственно участвует в синтезе хлорофилла, поскольку их дефицит вызывает обесцвечивание, или хлороз, у растений. Пиррольные кольца хлорофилла образуются в результате предварительного соединения глицина с янтарной кислотой. Между реакцией этих двух соединений и окончательным образованием комплексного соединения хлорофилла происходит ряд промежуточных реакций, катализируемых специфическими ферментами. Реакция между глицином и сукцинил-КоА зависит от присутствия железа в качестве кофермента, и дефицит этого элемента вызывает быстрый хлороз в молодых, развивающихся листьях. Дефицит марганца, калия, цинка и меди также вызывает хлороз, но роль этих элементов пока не изучена. Предположительно, они активируют определенные ферменты, участвующие в синтезе хлорофилла. Дефицит марганца приводит к разрушению внутренней ламеллярной структуры хлоропластов и может быть причиной ингибирования синтеза хлорофилла, стабилизирующего ламеллы.
Однако недостаток азота и магния вызывает пожелтение растений. Роль этих элементов понятна, поскольку они являются основными структурными компонентами хлорофилла. Это не исключает их более косвенных функций, например, магний, вероятно, действует как кофермент на одном или нескольких этапах синтеза хлорофилла.
4. Структура и синтез каротиноидных пигментов.
Каротиноиды — это группа пигментов, которые у высших растений обычно имеют красный, оранжевый, желтый или коричневый цвет и находятся в хлоропластах в сочетании с хлорофиллом. Иногда они также встречаются в других пластидах, например, в хромопластах корней моркови и томата, а также в хромопластах многочисленных желтых цветков. Желтый цвет каротиноидов виден у покрытосеменных растений, растущих в темноте и лишенных хлорофилла (что указывает на то, что свет не является необходимым для их накопления), а также осенью в листьях многих видов, где хлорофилл исчезает. Действительно, каротиноиды могут также иметь другие цвета, например, зеленый у авокадо, розовый у креветок или почти черный у головоногих моллюсков. Эти пигменты встречаются во всех основных группах растений и животных, но наше обсуждение будет сосредоточено в основном на тех, которые находятся в хлоропластах в сочетании с хлорофиллом. Каротиноиды можно разделить на две химические группы: каротины и ксантофиллы. В каждой группе существует несколько различных типов этих соединений, молекулы которых состоят из 40 атомов углерода. Каротиноиды, наиболее важным представителем которых является β-каротин, представляют собой углеводороды (рис. 2). Ксантофиллы, такие как лютеин и зеаксантин, имеют атомы кислорода, присоединенные к их концевым кольцам. И ксантофиллы, и каротины синтезируются в растениях из активного ацетата, т.е. ацетил-КоА, через мевалоновую кислоту.
Рис. 2. Структурные формулы каротиноидов.
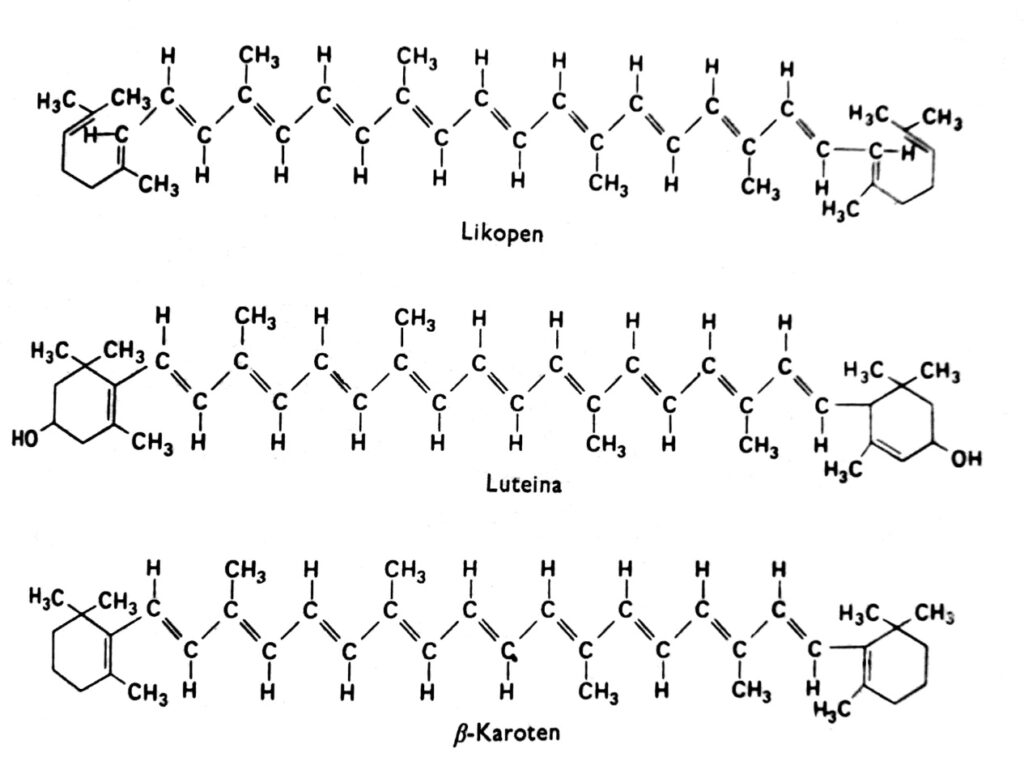
Практически ни один из каротиноидов не растворяется в воде, но они растворяются во многих органических растворителях, таких как ацетон и эфир. Ксантофиллы, содержащие дополнительные атомы кислорода, более гидрофильны, чем каротины. В растворе большинство каротиноидов поглощают преимущественно фиолетовый и синий свет в диапазоне 400-500 нм. Их спектры поглощения зависят от степени насыщенности двойных связей и, следовательно, могут немного отличаться в интактных хлоропластах. Есть данные, свидетельствующие о том, что в интактных клетках максимумы поглощения некоторых каротиноидов при 500 нм смещаются до 450 нм после экстракции этих пигментов органическим растворителем.
5. Функции каротиноидов.
Каротиноиды играют важную роль в фотосинтезе, вероятно, по двум причинам:
1. Они выполняют защитную функцию, оберегая хлорофилл от разрушительного воздействия света
и кислорода. Это разрушительное явление называется фотоокислением. Часто наблюдается обесцвечивание спиртового раствора хлорофилла под воздействием света, что сопровождается потреблением кислорода. Предположительно, подобная реакция обычно происходит в растениях, если каротиноиды не выполняют свою защитную функцию. Наиболее убедительные доказательства получены в результате наблюдений за некоторыми бактериальными мутантами. Поскольку они не содержат каротиноидных пигментов, фотоокисление в них происходит очень легко. Кроме того, известны мутанты подсолнечника-альбиносы, которые утратили способность синтезировать каротиноиды, но все еще производят следовые количества хлорофилла. В этом случае увеличение интенсивности света запускает дополнительное фотоокисление хлорофилла, и скорость этого процесса равна его синтезу. Механизм, посредством которого каротиноиды защищают хлорофилл от окисления, остается неясным. Однако считается, что каротиноиды фотоокисляются под воздействием света, который может разлагать хлорофилл.
2. Еще одна вероятная функция каротиноидов, и особенно ксантофиллов, заключается в их участии в поглощении света на длинах волн, активных в фотосинтезе. Существуют неопровержимые доказательства того, что некоторые каротиноиды, такие как фукоксантин (также называемый фукоксантолом), содержащийся в бурых водорослях (например, фукусе), активно участвуют в фотосинтезе. Эти доказательства были получены путем сравнения спектра поглощения фукоксантина с так называемым функциональным спектром, или спектром фотосинтетической активности. Фотосинтетический функциональный спектр — это график, иллюстрирующий эффективность фотосинтеза в зависимости от длин волн света, которые запускают этот процесс. Было обнаружено, что у зеленых растений и высших растений свет, поглощаемый некоторыми каротиноидами, также используется в фотосинтезе. В таких случаях максимумы функционального спектра попадают в диапазоны красного и синего света, где хлорофилл поглощает свет наиболее сильно. Однако фотосинтез по-прежнему происходит с использованием сине-зеленого света, предположительно потому, что некоторые каротиноиды активны в этом диапазоне. Наиболее убедительным доказательством участия каротиноидов в фотосинтезе является их способность передавать возбужденную энергию непосредственно хлорофиллу а. Когда растение облучают светом с длинами волн, поглощаемыми почти исключительно каротиноидами, наблюдается красная флуоресценция хлорофилла а. Предполагается, что энергия возбуждения, иногда высвобождаемая в виде флуоресценции, обычно используется в качестве движущей силы фотосинтеза.
6. Фикобилины.
Последняя группа фотосинтетических пигментов — фикобилины, обнаруженные в цианобактериях
и красных водорослях. Эти пигменты классифицируются на две группы: красные (фикоэритрины) и синие (фикоцианины). Как и хлорофилл, они состоят из четырех пиррольных колец. Однако фикобилины содержат открытую систему пиррольных колец и не содержат ни магния, ни фитольного фрагмента. В живых клетках они встречаются в ассоциации с белками и водорастворимы. Оба типа пигментов поглощают свет, используя его для фотосинтеза. У красных водорослей наибольшая эффективность фотосинтеза наблюдается в диапазоне зеленого света, поскольку в этом диапазоне фикоэритрин, в отличие от хлорофилла, поглощает свет наиболее интенсивно. Подобно каротиноидам, фикобилины передают возбужденную энергию хлорофиллу а и могут вызывать его флуоресценцию. Такой способ передачи энергии указывает на то, что фикобилины активно участвуют в фотосинтезе, захватывая фотоны, энергия которых обычно используется хлорофиллом а в качестве движущей силы фотосинтеза. Пигменты, такие как каротиноиды и фикобилины, которые передают энергию подходящему акцептору, например, хлорофиллу а, называются вспомогательными пигментами. В группу вспомогательных пигментов также входят хлорофиллы b, c и d, поскольку они поглощают световую энергию, которая, в свою очередь, может передаваться хлорофиллу а.
Литература:
-Франк Б. Салибери, Клеон Росс «Физиология растений», PWRiL, 1975.
-Отис Ф. Дэниел Г. Кертис Кларк «Введение в физиологию растений» PWRiL 1958.
-Зуржицкий Я. Мичневич М. «Физиология растений» PWRiL 1979.
— Рихтер Г. «Метаболические процессы в растениях», PWN, 1975.